|
|
| (4人の利用者による、間の35版が非表示) |
| 1行目: |
1行目: |
| <div align="right">
| | 両眼視差エネルギーモデル |
| <font size="+1">[http://researchmap.jp/rtanaka 田中 宏喜]</font><br>
| |
| ''京都産業大学 コンピュータ理工学部 インテリジェントシステム学科''<br>
| |
| DOI:<selfdoi /> 原稿受付日:2012年7月31日 原稿完成日:2012年8月13日<br>
| |
| 担当編集委員:[http://researchmap.jp/ichirofujita 藤田 一郎](大阪大学 大学院生命機能研究科)<br>
| |
| </div>
| |
|
| |
|
| 英:disparity energy model
| | 生体は両眼視差を手がかりに奥行きを知覚できるが、脳視覚系で左右眼からの入力が最初に収斂する第一次視覚野(V1野)には両眼視差の検出に理想化した神経細胞が存在する。この細胞の受容野機構を記述したモデルが視差エネルギーモデルである。 |
|
| |
|
| 同義語: binocular energy model
| | <br> |
| | |
| {{box|text=
| |
| 奥行きや物体の立体構造を知るための手がかりである[[両眼視差]]は、[[網膜]]からの視覚経路において[[第一次視覚野]](V1野)ではじめて検出される。この両眼視差(正確には絶対視差)の検出に特化したV1野細胞の[[受容野]]モデルが視差エネルギーモデルであり、現在、脳における両眼視差検出機構の標準モデルとなっている。1990年[[wikipedia:ja:カリフォルニア大学バークレー校|カリフォルニア大学バークレー校]]の大澤五住らによって提案された<ref name="ref1"><pubmed> 2396096 </pubmed></ref>。 以降、視差エネルギーモデルを出発点にして、相対視差など様々なタイプの両眼視差を検出する神経機構や、両眼対応点問題を解決するための脳内機構を調べる研究が数多く行われている。
| |
| }}
| |
|
| |
|
| == 両眼視差 == | | == 両眼視差 == |
|
| |
|
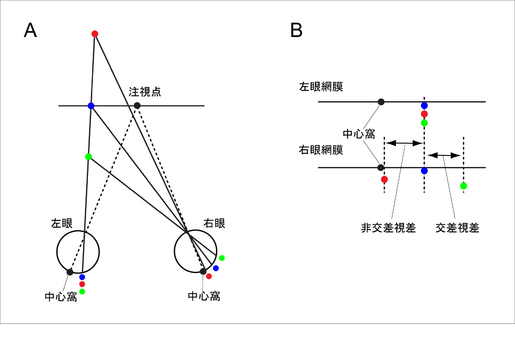
| [[Image:BinocularDisparity.png|thumb|350px|<b>図1 両眼視差</b><br />両眼視差. A,2つの眼で黒丸を注視するとき、さまざまな奥行きにある刺激の網膜投影像。B, 左右の網膜を平にして、上下に並べたもの。注視している点(注視点という)に位置する黒丸の像は、網膜で視力の最も高い場所である中心窩に投影される。注視点と同じ奥行きにある刺激(青)の左右の像は、中心窩を基準とした網膜座標上の同じ位置に投影され、その両眼視差はゼロとなる。一方、注視点と異なる奥行き面上にある刺激(赤、緑)は、左右網膜の異なる位置に投影され、ゼロ以外の両眼視差をもつ。手前にある刺激(緑)と、奥にある刺激(赤)の両眼視差の方向は逆になり、前者を交差視差、後者を非交差視差とよんでいる。]]
| | 物体が左右眼に投影されるとき、注視面上にある刺激(図1、青丸)の左右の像は、網膜中心窩を基準とした座標系の同じ位置に投影されるが、注視面と異なる奥行き面上にある物体は、網膜座標系の異なる位置に投影される。この網膜像の位置のずれのことを両眼視差という。両眼視差の量は刺激と注視面の奥行き距離に比例する。また刺激が注視点より手前にある場合(緑丸)と、奥にある場合(赤丸)とで、両眼視差の方向(符号)は逆になる。手前にある刺激により生じる両眼視差を交差視差、奥にある刺激により生じる両眼視差を非交差視差とよぶ。 |
|
| |
|
| われわれが両眼でものをみるとき、2つの眼が注視している点(注視点)と同じ奥行きにある刺激は、左右の網膜上の同じ位置に投影される(=いずれの網膜においても、網膜の中心である中心窩から同じ方向、量だけ離れた位置に投影される)のに対し、注視点と異なる奥行きにある刺激は水平方向にずれた位置に投影される(図1参照)。この網膜像の位置のずれのことを両眼視差という(単に視差ともいう)。両眼視差の量は刺激と注視点の奥行き距離に比例する。また刺激が注視点より手前にある場合と、奥にある場合とで両眼視差の方向(符号)は逆になる。慣習上、前者にはマイナス、後者にはプラスの符号をつけ、前者を交差視差 、後者を非交差視差とよぶ。
| | <br> |
|
| |
|
| われわれが両眼視差のみを手がかりにして奥行きを知覚できることは1838年にチャールズ・ホイートストン([[wikipedia:Charles Wheatstone|Wheatstone]])がハプロスコープ(ミラー式両眼視システム)を考案して証明した。
| | [[Image:BinocularDisparity.png|515x366px|<b>図1 両眼視差</b> 左の図は、左右の眼で点Fを注視しているとき、注視点および異なる奥行き面にある3つの点(赤、青、緑の丸)が網膜に投影される様子を上から見た断面図である。右の図は、この断面図のうち、左右の網膜投影像をまっすぐ伸ばして、上下に並べたものである。注視点Fは、左右眼の光軸(左の図の点線)をとおり、網膜で視力の最も高い中心窩(0で表す)に投影される。 注視点と同じ奥行きにある面、すなわち注視面上にある刺激(青丸)の左右の像は、中心窩を基準とした網膜座標上の同じ位置に投影される。両眼視差とは、この座標系における左右の網膜像の位置のずれのことである。したがって、青丸刺激の両眼視差はゼロである。注視面と異なる奥行き面上にある刺激は、左右網膜の異なる位置に投影されるので、ゼロ以外の両眼視差をもつ。両眼視差の量は、刺激の注視面からの距離に比例する。また、手前にある刺激(緑丸)では左眼像に対し右眼像が右側に、奥にある刺激(赤丸)では左眼像にたいし右眼像が左側にずれるため、注視点の手前にある刺激と、奥にある刺激とでは両眼視差の符号が逆になる。慣習により、手前にある刺激がもつ両眼視差にはマイナスの符号をつけ、奥にある刺激がもつ両眼視差にはプラスの符号をつける。また前者を交差視差、後者を非交差視差とよんで区別している。]] |
|
| |
|
| == V1野にみられる両眼視差選択性 == | | == V1野にみられる両眼視差選択性 == |
|
| |
|
| 網膜からの視覚処理経路において、左右両眼に受容野をもつ細胞はV1野で初めて現れる<ref name="ref2"><pubmed> 14403679 </pubmed></ref>。これらの細胞の一部は、刺激の両眼視差がある範囲に入るときには強く応答し、それ以外のときにはあまり応答しない特性、すなわち両眼視差選択性を示す<ref name="ref4"><pubmed> 6065881 </pubmed></ref>。ゼロ視差やさまざまな大きさの交差視差、非交差視差を最適とする細胞が存在する。初期の研究ではこれらの細胞は6つのタイプに分類されたが<ref name="ref7"><pubmed> 3199191 </pubmed></ref>、最近の研究結果は、これらは1つの連続体として捉えたほうがよいことを示している <ref name="ref8"><pubmed> 11784742 </pubmed></ref>。
| | 網膜からの視覚処理経路において、左右眼に受容野をもつ両眼性の細胞が、第一次視覚野(V1野)に初めて現れる。受容野構造の違いから、V1野の細胞は単純型細胞と複雑型細胞とに大きく分類されるが、いずれのタイプでも両眼性の細胞は存在する。うち一部は、視覚刺激がある両眼視差をもつときには強く応答し、それ以外のときにはあまり応答しない特性、すなわち、両眼視差選択性を示す。両眼視差ゼロを最適とする細胞や、さまざまな大きさの交差視差、非交差視差に選択性をもつ細胞が存在している。<br> 単純型細胞の両眼視差選択性は、視覚刺激の場所やコントラストに依存する。一方、複雑型細胞の両眼視差選択性はそれらに依存せず一定である。したがって、複雑型細胞のほうが両眼視差を検出するうえで理想的な振る舞いをしているといえる。このような複雑型細胞の両眼視差選択性を作り出す受容野内部機構として提唱されたモデルが、両眼視差エネルギーモデルである。このモデルにおいて、複雑型細胞の出力は、両眼性単純型細胞フィードフォワード結合で表される。以下に両眼性単純型細胞の受容野構造および両眼視差エネルギーモデルを説明する。 |
| | |
| 受容野構造の違いから、V1野の細胞は[[単純型細胞]]と[[複雑型細胞]]とに大きく分類され、いずれのタイプにも両眼視差選択性細胞は存在する。しかし、後述するように、単純型細胞よりも複雑型細胞のほうが両眼視差を検出するうえで理想的な振る舞いをする。この複雑型細胞の両眼視差選択性を説明する受容野モデルが視差エネルギーモデルである。このモデルで複雑型細胞の応答は、複数の単純型細胞からの入力の和で生成される。以下に単純型細胞の両眼受容野構造を述べ、続いて視差エネルギーモデルを説明する。
| |
|
| |
|
| == 単純型細胞の受容野構造と両眼視差選択性 == | | == 単純型細胞の受容野構造と両眼視差選択性 == |
|
| |
|
| [[Image:BinocularSimple.png|thumb|350px|<b>図2 単純型細胞の受容野構造と両眼視差選択性</b><br />A. 単純型細胞の両眼受容野構造. 左右の受容野のx-y2次元構造とx-方向の1次元断面図を示している。これらの受容野を持つ細胞を丸で表し、その下の図で半波整流機構を表している。B-D. 単純型細胞の視差選択性。受容野の上の四角は、明るいスポット刺激の左右網膜像を表し、この細胞が最大応答を示すように配置されている。B. ゼロ視差を最適とする受容野構造. C. 位置モデルにより非交差視差に適刺激とする受容野。D. 位相モデルにより非交差視差を適刺激とする受容野。<br />]] | | [[Image:BinocularSimple.png|400 px|'''図2 単純型細胞の受容野構造'''<br/>]] |
| | |
| 単純型細胞細胞は、明るい刺激に応答するON領域と暗い刺激に応答するOFF領域が分離した受容野をもつ。受容野の空間構造は[[wikipedia:ja:ガボール関数|ガボール関数]]で近似できる。
| |
|
| |
|
| 多くの単純型細胞は両眼に受容野をもつ。これら両眼性単純型細胞の応答は、両眼からの入力を左右の受容野で重みづけをして足し合わせたのち、さらに[[wikipedia:ja:半波整流|半波整流]]をしたものとして表すことができる(図2A)。
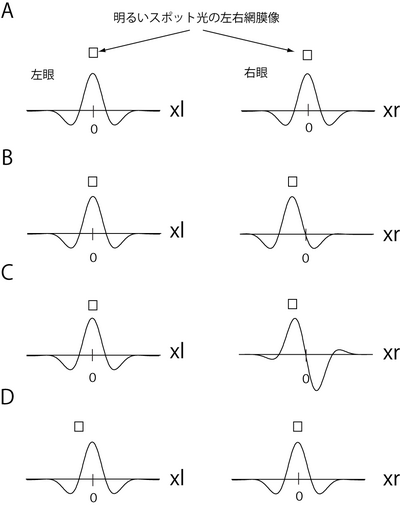
| | 単純型細胞の受容野では、明るい刺激に応答するON領域と暗い刺激に応答するOFF領域が分離しており、その空間構造はガボールフィルターで近似できる。多くの細胞が両眼性であり、その応答は、両眼からの信号をそれぞれ左右の受容野で重みづけして線形加算したのち、半端整流したものとして記述される(図2)。<br> このような受容野をもつ単純型細胞の両眼視差選択性は、大きく分けて2つの機構で生じることが知られている。1つは、同じ空間構造の受容野の位置が左右の眼でずれることで、細胞がそのずれと等しい両眼視差を最適とする機構であり、「位置モデル」とよばれている。たとえば、左右の受容野に位置のずれがない図3Aの細胞は、刺激の両眼視差がゼロのとき最も強く応答するのにたいし、左右の受容野の位置がずれた図3Bの細胞は、刺激の左右像が受容野のずれとマッチした両眼視差をもつとき、最も強く応答する。2つ目の機構は、受容野の中心位置は同じであるが、受容野の(ガボール)位相が左右で異なる(図3C)ことで、細胞が両眼視差に選択性をもつ機構で、この機構は「位相モデル」とよばれている。<br> 単純型細胞の多くは両眼視差に依存した応答を示す。ただし、単純型細胞の両眼視差依存性は、刺激の左右投影像の単眼上での位置や、刺激のコントラストにも大きく依存するという問題がある。たとえば、図3Dのように、明るいスポット光の左眼像の位置を、受容野の中心よりもやや左に固定する場合、左右に同じ受容野をもつ細胞が最もよく反応する視差は、ゼロ視差ではなく交差視差となる。このような問題のため、通常、単純型細胞が第一次視覚野の両眼視差検出器として取り扱われることはない。<br> |
|
| |
|
| 単純型細胞が視差選択性をもつ機構は、大きく分けて2種類ある<ref name="ref9"><pubmed> 2067576 </pubmed></ref><ref name="ref12"><pubmed> 11784743 </pubmed></ref>。1つは、左右の受容野の形は同じであるが、その位置がずれることにより視差選択性が生じる機構で「位置モデル」と呼ばれている。細胞は受容野の位置のずれと等しい両眼視差に最も強く応答する。たとえば、図2Bの受容野をもつ細胞はゼロ視差に最も強く応答し、図2Cの受容野をもつ細胞は非交差視差に最も強く応答する。第2の機構は、受容野の(中心)位置は同じであるが、受容野の形(位相)が異なることにより両眼視差選択性が生じる機構で、「位相モデル」とよばれている(図2D)。さらに実際の細胞には、位置と位相の両方ずれた受容野をもつものも多く、このような受容野に基づく視差選択性機構は「ハイブリッドモデル」とよばれている。様々な機構がある意義については、これら3種類の機構を利用することで対応点問題(後述)がより精度よく解けることが理論的に示されている<ref name="read"><pubmed> 17828262 </pubmed></ref>。
| | [[Image:PositionPhase.png|400 px|'''図3 単純型細胞受容野と両眼視差選択性'''<br/>]] |
|
| |
|
| 多くの両眼性単純型細胞は、両眼視差によって大きな応答の変動を示す。しかし細胞が最大応答する両眼視差は、刺激の左右投影像の単眼上での位置や、刺激のコントラストにも依存するという問題がある。このような問題のため、通常、単純型細胞がV1野の両眼視差検出器のモデルとして取り扱われることはない。
| |
|
| |
|
| == 視差エネルギーモデル == | | == 視差エネルギーモデル == |
|
| |
|
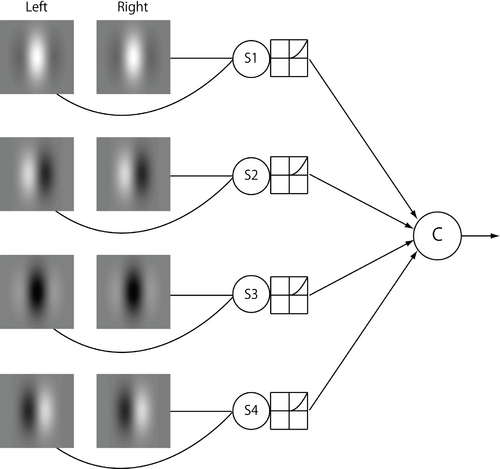
| [[Image:DisparityEnergyModel.png|thumb|350px|<b>図3 視差エネルギーモデル</b><br />複雑型細胞を模倣したエネルギーユニット(Cの記号で表す)は、両眼性単純型細胞を模倣した4つのサブブユニット(S1, S2, S3, S4)が出す信号を線形加算し、外部に出力する。詳細は本文参照。<br />]]
| | 単純型細胞の両眼視差選択性は、視覚刺激の(単眼)位置やコントラストに依存するのにたいし、複雑型細胞の両眼視差選択性はそれらに依存せず一定である。このような複雑型細胞の両眼視差選択性を作り出す受容野内部機構として提唱されたモデルが、視差エネルギーモデルであり、図4のように表される。このモデルにおいて、複雑型細胞(Cの記号で表す)は、両眼性単純型細胞をモデル化した4つのサブブユニット(S1, S2, S3, S4)が出す信号を線形加算し、外部に出力する。4つのサブユニットのガボールフィルターの位相は、右眼、左眼のそれぞれにおいて90度ずつ異なっている。また各サブニットにおいて、左右ガボールフィルターの両眼間の位相差は同一である。この両眼位相差を(4つのサブユニットで同一に保ちながら)変化させることで、モデルの両眼視差選択性を変化させることができる。<br> 刺激の左右の像が、複雑型細胞の最適な両眼視差をもつ場合(図3の場合はゼロ視差)、受容野内部のどの場所に刺激がくる場合でも、4つのサブユニットのいずれかが強く応答する。図3の場合、明るい刺激が受容野内部の中心付近に呈示される場合にはS1が、左部分に呈示される場合にはS2が、右部分に呈示される場合にはS4がそれぞれゼロ視差に強く応答する。また、背景より暗い刺激が受容野の中心付近、右部分、左部分に呈示される場合には、S4、S3、S2がそれぞれゼロ視差に強く応答する。このため、複雑型細胞は、受容野内部の刺激の位置やコントラストに影響されずに、同じ両眼視差選択性を示すようになり、両眼視差の検出器としては理想的な振る舞いをする。<br> |
| | |
| 単純型細胞の両眼視差選択性が視覚刺激の単眼位置やコントラストに依存するのに対し、複雑型細胞の両眼視差選択性はそれらに依存せず一定となる。この複雑型細胞の特性を説明するモデルが視差エネルギーモデルであり、図3のように表される<ref name="ref1" /><ref name="ref13"><pubmed> 9212245 </pubmed></ref>。このモデルにおいて、複雑型細胞(Cの記号で表す)は、両眼性単純型細胞をモデル化した4つのサブユニット(S1, S2, S3, S4)が出す信号を[[wikipedia:ja:線形加算|線形加算]]し、外部に出力する。4つのサブユニットの受容野の位相は、右眼、左眼のそれぞれにおいて90度ずつ異なっている。サブユニットの左右受容野の方位、空間周波数は全て同じである。
| |
| | |
| 各サブニットにおいて、左右受容野の両眼間の[[wikipedia:ja:位相|位相]]差を(4つのサブユニットで同一に保ちながら)変化させることで、モデルの両眼視差選択性を変化させることができる。あるいは、両眼間位相差を0にしたまま、受容野の位置を両眼間で一定量ずらすことでも、モデルの両眼視差選択性を変化させうる。前者は単純型細胞の「位相モデル」に対応し、後者は「位置モデル」と対応する。図3の例では、サブユニットの左右の受容野は位相、位置ともにずれがなく、モデルはゼロ視差に選択性をもつ。<br> 刺激の左右の像が、複雑型細胞の最適な両眼視差をもつ場合、受容野内部のどの場所に刺激がくる場合でも、4つのサブユニットのいずれかが強く応答する。図3の場合、明るいゼロ視差の刺激が受容野の中心に呈示される場合にはS1が、左部分に呈示される場合にはS2が、右部分に呈示される場合にはS4がそれぞれ強く応答する。また、背景より暗いゼロ視差の刺激が受容野の中心、左部分、右部分に呈示される場合には、S3、S4、S2がそれぞれ強く応答する。このため、複雑型細胞は、受容野内部の刺激の位置やコントラストに影響されずに、同じ両眼視差選択性を示すようになり、両眼視差の検出器としては理想的な振る舞いをする。
| |
| | |
| 視差エネルギーモデルは、最小4つのサブユニットの組み合わせで複雑型細胞の特性を表しうることを述べたものであり、複雑型細胞が必ず4つの単純型細胞の入力により生成されることを提唱しているわけではない。実際には、4つ以上の単純型細胞の入力により複雑型細胞の受容野は形成されていると推定されている<ref name="sasaki"><pubmed> 20943923</pubmed></ref> 。<br>
| |
| | |
| 視差エネルギーモデルが行っている計算は、2枚の画像についての局所的な相関計算と類似性がある。2枚の画像の局所相関を計算する場合、2枚の画像を一定量ずらしたとき同じ位置にくる画素値をかけあわせて、その局所平均をとる。視差エネルギーモデルが行っていることは数学的にはこのような計算と捉えることができる<ref name="qian"><pubmed> 9274767</pubmed></ref> 。
| |
| | |
| == 視差エネルギーモデルの拡張による種々の両眼視差の検出 ==
| |
| | |
| === 相対視差 ===
| |
| | |
| ここまで扱ってきた両眼視差は、[[中心窩]]を基準とした座標系における、左右網膜像の位置のずれとして定義されたものであり、絶対視差とも呼ばれる。これに対して、2つの刺激がもつ絶対視差の差異のことを相対視差と呼ぶ。われわれは隣接する刺激の奥行きを非常に精度よく弁別できるが、これには[[眼球]]の[[輻輳運動]]の影響をうけない相対視差が利用されていると考えられている。[[wikipedia:ja:サル|サル]]V1野の細胞の大部分は絶対視差をコードしているが、V1野から入力を受ける[[wikipedia:ja:V2野|V2野]]や[[wikipedia:ja:V4野|V4野]]には、相対視差に選択的応答を示す細胞が一定の割合で存在する<ref name="ref14"><pubmed> 11967544 </pubmed></ref><ref name="ref15"><pubmed> 17507498 </pubmed></ref>。この選択性は、異なる場所に受容野をもち、異なる絶対視差に選択性をもつ視差エネルギーモデルの出力を2段階的に統合することで生成できる。<ref name="ref14" />。
| |
| | |
| === 視差の勾配 ===
| |
| | |
| 前額平行面から奥行き方向に傾いた物体表面をみるとき、その表面上の特徴点は網膜上で両眼視差の勾配を作り出す。受容野内部の場所によって、異なる視差選択性をもつサブユニットから入力を受け取るように、視差エネルギーモデルを拡張すると(このときサブユニットの数は4よりも多くなる)、この拡張モデルは視差勾配への選択性をもつようになる(ただしこの選択性は絶対視差にも強く依存する)。[[wikipedia:ja:ネコ|ネコ]][[V1野]]にはこのモデルに合致するほうな細胞が存在することが示されている<ref name="sanada"><pubmed> 16394073</pubmed></ref>。
| |
| | |
| === 方位視差 ===
| |
| | |
| 前額平行面から奥行き方向に傾いた線分が網膜に投影されるとき、その左右の投影像の方位にはずれが生じる。この方位視差を手がかりに、われわれは線分の奥行き方向の傾きを知ることができる。4つのサブユニットにおいて、左右のガボールフィルターの方位が同じ角度だけずれることを許容すると、視差エネルギーモデルは方位視差に対する感受性を持つようになり、サルV1野には実際にこのような特性を示す細胞が報告されている。さらに、様々な受容野方位をもつサブニットからの入力を集めるようにモデルを拡張すると、この拡張モデルは、刺激の左右の網膜像の方位自身には依存しない方位視差選択性を示すようになるが、そのような特性を示す細胞は報告されていない<ref name="bridge"><pubmed> 11549739 </pubmed></ref>。
| |
| | |
| === 二次特徴の両眼視差 ===
| |
| | |
| 視覚系が利用可能な両眼視差のうち、最も強い奥行き手がかりとなるものは輝度エッジから生じる両眼視差である。しかし、テクスチャーエッジ(例 : 縦縞模様の領域と横縞模様の領域の境界)など二次特徴とよばれる刺激により生じる両眼視差からも奥行き知覚は可能である。視覚野の細胞の多くは輝度エッジの両眼視差にしか応答しない。しかしながら、ネコ[[18野]](細胞構築学的にはV2野とされる)の一部の細胞は、二次特徴の両眼視差に選択性をもつことが示されている。このような細胞は、輝度のエッジにも同じ両眼視差に最大応答し、特徴手がかりに依存しない両眼視差選択性を示す。2次特徴の両眼視差は、両眼視差エネルギーモデルの各サブユニットの左右受容野を、線形フィルターではなく『フィルター>整流>フィルター』というカスケード型の非線形機構で置き換えることで検出できる<ref name="ref16"><pubmed> 16624957 </pubmed></ref>。
| |
| | |
| == 視差エネルギーモデルと両眼対応点問題 ==
| |
| | |
| 両眼視差を正しく検出するためには、左眼の網膜像のどの特徴と右眼の網膜像のどの特徴とが対応するのか(同じ外界刺激の投影像であるのか)を正しく決めることが不可欠である。この課題を両眼対応点問題とよぶ。刺激が視野の中にただ1つしか存在せず、左右の網膜上にはその投影像が1つずつしか存在しない状況では解は自明である。しかし、視野の中に似た刺激が多数存在し、左右の網膜上に似た特徴が多数存在する状況下では、この対応づけは容易ではない。
| |
| | |
| 上記の多数の刺激が存在する状況では、正しくない組み合わせ(=フォールスマッチ)が細胞の左右の受容野内部に入る状況は頻繁に起こる。このとき視差エネルギーモデルはフォールスマッチにも応答することが示されている。しかしながら、われわれの視覚系は、フォールスマッチに基づいて誤った奥行きを知覚することはなく、正しい組み合わせ(=コレクトマッチ)に基づいて奥行きを知覚している。このためには視差エネルギーモデルが出力するフォールスマッチの信号を遮断し、コレクトマッチの信号を選び出す神経機構が必要となる。
| |
| | |
| V1野細胞は、視差エネルギーモデルの予測よりは低いものの、フォールスマッチにも強く応答する<ref name="ref17"><pubmed> 9212245 </pubmed></ref><ref name="ref18"><pubmed> 9305841 </pubmed></ref><ref name="ref19"><pubmed> 10844045 </pubmed></ref>。一方でサルV4野や[[IT野]]など腹側視覚経路の細胞はフォールスマッチにはあまり応答しない<ref name="ref20"><pubmed> 15371518 </pubmed></ref><ref name="ref21"><pubmed> 12597865 </pubmed></ref>。このことは視差情報がこの経路に沿って処理されるなかで、対応点問題が解決されていることを示している。対応点問題を解決するための神経機構としては、空間周波数チャネルの収斂に基づく機構や<ref name="ref20"><pubmed> 8759452 </pubmed></ref>、位置モデル、位相モデル、ハイブリッドモデルが出す視差信号全体を利用した機構などが提案されている<ref name="read" />。V4野では周波数チャネルの収斂が実際に起こっていることが示されている<ref name="kumano"><pubmed> 17959744 </pubmed></ref>。
| |
| | |
| V1野複雑型細胞の応答は、基本的には視差エネルギーモデルでよく説明できるが、前述したようにフォールスマッチへの応答が視差エネルギーモデルの予測より減弱する。さらに視差エネルギーモデルの予測より、自然界に実在する両眼視差のパターンにたいしてより大きな応答変動をすることも示されている。このような応答を説明する機構の1つして、複雑型細胞が4つ以上の単純型細胞から興奮および抑制入力を受けるモデルが提案されている<ref name="haefnar"><pubmed> 18184571</pubmed></ref> <ref name="tanabe2011"><pubmed> 21632950</pubmed></ref>。
| |
|
| |
|
| == 参考文献 ==
| | [[Image:DisparityEnergyModel.png|500 px|'''図4 視差エネルギーモデル'''<br/>]] |
|
| |
|
| <references />
| | <br> |
両眼視差エネルギーモデル
生体は両眼視差を手がかりに奥行きを知覚できるが、脳視覚系で左右眼からの入力が最初に収斂する第一次視覚野(V1野)には両眼視差の検出に理想化した神経細胞が存在する。この細胞の受容野機構を記述したモデルが視差エネルギーモデルである。
両眼視差
物体が左右眼に投影されるとき、注視面上にある刺激(図1、青丸)の左右の像は、網膜中心窩を基準とした座標系の同じ位置に投影されるが、注視面と異なる奥行き面上にある物体は、網膜座標系の異なる位置に投影される。この網膜像の位置のずれのことを両眼視差という。両眼視差の量は刺激と注視面の奥行き距離に比例する。また刺激が注視点より手前にある場合(緑丸)と、奥にある場合(赤丸)とで、両眼視差の方向(符号)は逆になる。手前にある刺激により生じる両眼視差を交差視差、奥にある刺激により生じる両眼視差を非交差視差とよぶ。
V1野にみられる両眼視差選択性
網膜からの視覚処理経路において、左右眼に受容野をもつ両眼性の細胞が、第一次視覚野(V1野)に初めて現れる。受容野構造の違いから、V1野の細胞は単純型細胞と複雑型細胞とに大きく分類されるが、いずれのタイプでも両眼性の細胞は存在する。うち一部は、視覚刺激がある両眼視差をもつときには強く応答し、それ以外のときにはあまり応答しない特性、すなわち、両眼視差選択性を示す。両眼視差ゼロを最適とする細胞や、さまざまな大きさの交差視差、非交差視差に選択性をもつ細胞が存在している。
単純型細胞の両眼視差選択性は、視覚刺激の場所やコントラストに依存する。一方、複雑型細胞の両眼視差選択性はそれらに依存せず一定である。したがって、複雑型細胞のほうが両眼視差を検出するうえで理想的な振る舞いをしているといえる。このような複雑型細胞の両眼視差選択性を作り出す受容野内部機構として提唱されたモデルが、両眼視差エネルギーモデルである。このモデルにおいて、複雑型細胞の出力は、両眼性単純型細胞フィードフォワード結合で表される。以下に両眼性単純型細胞の受容野構造および両眼視差エネルギーモデルを説明する。
単純型細胞の受容野構造と両眼視差選択性
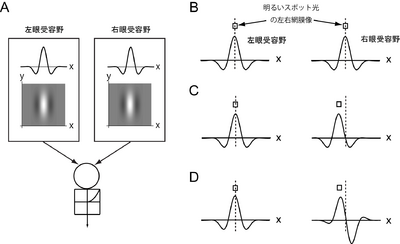
単純型細胞の受容野では、明るい刺激に応答するON領域と暗い刺激に応答するOFF領域が分離しており、その空間構造はガボールフィルターで近似できる。多くの細胞が両眼性であり、その応答は、両眼からの信号をそれぞれ左右の受容野で重みづけして線形加算したのち、半端整流したものとして記述される(図2)。
このような受容野をもつ単純型細胞の両眼視差選択性は、大きく分けて2つの機構で生じることが知られている。1つは、同じ空間構造の受容野の位置が左右の眼でずれることで、細胞がそのずれと等しい両眼視差を最適とする機構であり、「位置モデル」とよばれている。たとえば、左右の受容野に位置のずれがない図3Aの細胞は、刺激の両眼視差がゼロのとき最も強く応答するのにたいし、左右の受容野の位置がずれた図3Bの細胞は、刺激の左右像が受容野のずれとマッチした両眼視差をもつとき、最も強く応答する。2つ目の機構は、受容野の中心位置は同じであるが、受容野の(ガボール)位相が左右で異なる(図3C)ことで、細胞が両眼視差に選択性をもつ機構で、この機構は「位相モデル」とよばれている。
単純型細胞の多くは両眼視差に依存した応答を示す。ただし、単純型細胞の両眼視差依存性は、刺激の左右投影像の単眼上での位置や、刺激のコントラストにも大きく依存するという問題がある。たとえば、図3Dのように、明るいスポット光の左眼像の位置を、受容野の中心よりもやや左に固定する場合、左右に同じ受容野をもつ細胞が最もよく反応する視差は、ゼロ視差ではなく交差視差となる。このような問題のため、通常、単純型細胞が第一次視覚野の両眼視差検出器として取り扱われることはない。
視差エネルギーモデル
単純型細胞の両眼視差選択性は、視覚刺激の(単眼)位置やコントラストに依存するのにたいし、複雑型細胞の両眼視差選択性はそれらに依存せず一定である。このような複雑型細胞の両眼視差選択性を作り出す受容野内部機構として提唱されたモデルが、視差エネルギーモデルであり、図4のように表される。このモデルにおいて、複雑型細胞(Cの記号で表す)は、両眼性単純型細胞をモデル化した4つのサブブユニット(S1, S2, S3, S4)が出す信号を線形加算し、外部に出力する。4つのサブユニットのガボールフィルターの位相は、右眼、左眼のそれぞれにおいて90度ずつ異なっている。また各サブニットにおいて、左右ガボールフィルターの両眼間の位相差は同一である。この両眼位相差を(4つのサブユニットで同一に保ちながら)変化させることで、モデルの両眼視差選択性を変化させることができる。
刺激の左右の像が、複雑型細胞の最適な両眼視差をもつ場合(図3の場合はゼロ視差)、受容野内部のどの場所に刺激がくる場合でも、4つのサブユニットのいずれかが強く応答する。図3の場合、明るい刺激が受容野内部の中心付近に呈示される場合にはS1が、左部分に呈示される場合にはS2が、右部分に呈示される場合にはS4がそれぞれゼロ視差に強く応答する。また、背景より暗い刺激が受容野の中心付近、右部分、左部分に呈示される場合には、S4、S3、S2がそれぞれゼロ視差に強く応答する。このため、複雑型細胞は、受容野内部の刺激の位置やコントラストに影響されずに、同じ両眼視差選択性を示すようになり、両眼視差の検出器としては理想的な振る舞いをする。