|
|
| (4人の利用者による、間の80版が非表示) |
| 1行目: |
1行目: |
| <div align="right">
| | 英語1: Myelinated nerve 英語2: Medullated nerve |
| <font size="+1">[http://researchmap.jp/9190 清水 崇弘]</font><br>
| |
| ''University College London (Wolfson Institute for Biomedical Research)''<br>
| |
| <font size="+1">[http://researchmap.jp/kazuhiroikenaka 池中 一裕]</font><br>
| |
| ''自然科学研究機構 生理学研究所 分子生理研究系・分子神経生理''<br>
| |
| DOI:<selfdoi /> 原稿受付日:2012年8月29日 原稿完成日:2012年11月20日 原稿更新日:2013年8月21日<br>
| |
| 担当編集委員:[http://researchmap.jp/michisukeyuzaki 柚崎 通介](慶應義塾大学 医学部生理学)<br>
| |
| </div>
| |
|
| |
|
| 英語名: myelinated nerve, medullated nerve 独:myelinisierte Nervenfaser 仏:fibres myélinisées
| | 中枢神経系では希突起膠細胞(あるいはオリゴデンドロサイト[oligodendrocyte])、末梢神経系ではシュワン細胞(Schwann cell)というグリア細胞の形成する髄鞘(ミエリン)[参照:「髄鞘」[http://bsd.neuroinf.jp/w/index.php?title=%E9%AB%84%E9%9E%98&redirect=no]]が神経軸索[参照:「軸索」[[http://bsd.neuroinf.jp/w/index.php?title=%E8%BB%B8%E7%B4%A2&action=edit&redlink=1]]]の周囲に形成された神経のことをいう。神経軸索は樹状突起と比べると細く、長く、平滑な細胞突起である。ミエリン鞘は等間隔にあるランヴィエの絞輪(node of Ranvier)で途切れていて、軸索中のNa+チャネルはほぼこの節に集中している。 |
|
| |
|
| {{box|text=
| | 髄鞘はリン脂質に富んだタンパク質で層構造を形成しているため、髄鞘は絶縁体の役割を果たし、軸索膜を絶縁して膜からの電流のもれをほぼ完全に防ぎ、神経細胞からの電気信号を跳躍伝導させることができる。また、単に絶縁体としての働きだけではなく、神経軸索の保護や軸索との間に緊密な相互作用を行うことで、さまざまな神経機能を調節している。主要構成蛋白質はミエリン塩基性蛋白質(Myelin basic protein; MBP)、プロテオリピドプロテイン(myelin proteolipid protein; PLP)ミエリン関連糖タンパク質(Myelin associated glycoprotein; MAG)、ミエリンオリゴデンドロサイト糖タンパク質(Myelin oligodendrocyte glycoprotein; MOG)などがある。 <br> <br> |
| [[髄鞘]]([[ミエリン]])を有する[[軸索]]のことを有髄線維あるいは[[有髄神経]]線維という。髄鞘は[[中枢神経系]]では[[稀突起膠細胞]](あるいは[[オリゴデンドロサイト]][oligodendrocyte])、[[末梢神経系]]では[[シュワン細胞]](Schwann cell)という[[グリア細胞]]が形成する。軸索は樹状突起と比べると細く平滑な細胞突起である。[[ミエリン鞘]]は等間隔にある[[ランヴィエ絞輪]](node of Ranvier)によって区切られており、軸索中の[[電位依存性ナトリウムチャネル]]はランヴィエ絞輪に集積している。
| | == 1:神経の分類と髄鞘の有無 == |
| | 末梢神経の神経線維は髄鞘の有無、直径、伝導速度等で分類される。有髄線維と無髄線維では有髄線維が、同じ種類の線維間では直径が大きい方が伝導速度が速い。前者は跳躍伝導により、後者は電気緊張電位の広がりの違いにより速い速度を得ている。一般に骨格筋運動と付随する固有感覚、部位のはっきりした皮膚感覚は伝導速度の速い線維を、交感神経活動や鈍痛などは伝導速度の遅い線維を利用して伝えられる。 |
| | [[Image:神経太さ分類.png|RTENOTITLE]]<br> |
|
| |
|
| 髄鞘は[[リン脂質]]に富んだ[[wikipedia:ja:タンパク質|タンパク質]]で層構造を形成しているため、非常に良い[[絶縁体]]の役割を果たす。さらに、活動電位を発生させる電位依存性[[ナトリウムチャネル]]がランヴィエ絞輪に集積しているために、活動電位を軸索に沿って[[跳躍伝導]]させることができる。また、単に絶縁体としての働きだけではなく、軸索との間に物質交換を行うことによって神経軸索の栄養・保護などさまざまな神経機能を調節している。主要構成タンパク質は[[ミエリン塩基性タンパク質]]([[myelin basic protein]]; [[MBP]])、[[ミエリンプロテオリピドプロテイン]]([[myelin proteolipid protein]]; [[PLP]])であり、その他に[[ミエリン関連糖タンパク質]]([[myelin associated glycoprotein]]; [[MAG]])、[[ミエリンオリゴデンドロサイト糖タンパク質]]([[myelin oligodendrocyte glycoprotein]]; [[MOG]])、[[2',3'-環状ヌクレオチドホスホジエステラーゼ]]([[2',3'-cyclic-nucleotide phosphodiesterase]]; [[CNPase]])などを含む。
| | 図1:神経繊維の分類と髄鞘の有無 |
| }}
| |
|
| |
|
| == 神経の分類と髄鞘の有無 ==
| | <br> |
|
| |
|
| 末梢神経の神経線維は伝導速度によって大きくA,B,C線維と分類される。A線維はさらに速度の速い順にalpha,beta,gamma, deltaと分類される。例えばAalphaは固有知覚を求心性に伝えるとともに体性運動を遠心性に伝える。一方、感覚神経では太さに応じてI,II,III,IV線維と分類される。筋からの求心線維には慣用的にI-IVの分類が主に用いられ、皮膚からの求心線維にはA-Cの分類が用いられる。また、この伝導速度は髄鞘の有無や神経線維の直径の大小の組み合わせで規定される。同じ直径の有髄線維と[[無髄線維]]では有髄線維が、同じ種類の線維間では神経線維の直径の大きい方が伝導速度が速い。前者は跳躍伝導により、後者の[[電気緊張電位]]の広がりを利用した伝導よりも速い伝導速度を得ている。一般に[[wikipedia:ja:骨格筋|骨格筋]]運動と付随する[[固有感覚]]、部位のはっきりした[[皮膚感覚]]は伝導速度の速い線維を、[[交感神経]]活動や[[鈍痛]]などは伝導速度の遅い線維を利用して伝えられる。
| |
|
| |
|
| {| cellspacing="1" cellpadding="1" border="1"
| | == 2:有髄神経の構造 == |
| |+'''表1.神経線維の分類'''<ref name="ref1">'''Eric R. Kandel et al.'''<br>Principles of neural science fifth edition p475-480<br>''McGraw-Hill Professional, New York, 2012''</ref>
| | <br> 神経軸索の髄鞘と髄鞘の隙間は特別な名称がつけられており、非常に特異な構造をしている。ノード(node)に電位作動型イオンチャネルNav1.2や Nav1.6、アンキリンG(AnkG)などが、パラノード(paranode)にはCasprなど、ジャクスタパラノード(jaxtaparanode)にはKv1.2などのカリウムチャネルなどが分布する。中枢神経系(オリゴデンドロサイト)と末梢神経系(シュワン細胞)で髄鞘の巻き方が少し異なる。オリゴデンドロサイトは離れた軸索に別々に髄鞘を形成するのに対し、シュワン細胞はいくつかの軸索を抱え込むようにして包んだ後、1本の軸索を選別して、その周りに髄鞘を形成する。[]髄鞘を構成する蛋白質も少し異なる。 |
| |-
| |
| |
| |
| | style="text-align:center" | 筋からの求心性感覚神経線維(太さから分類)
| |
| | style="text-align:center" | 線維直径(μm)
| |
| | style="text-align:center" | 皮膚からの求心性感覚神経線維(速度から分類)
| |
| | style="text-align:center" | 伝導速度(m/s)
| |
| |-
| |
| | rowspan="4" style="text-align:center" | 有髄神経
| |
| | style="text-align:center" | I
| |
| | style="text-align:center" | 12-20
| |
| | style="text-align:center" | [[Aα]]
| |
| | style="text-align:center" | 72-120
| |
| |-
| |
| | style="text-align:center" | II
| |
| | style="text-align:center" | 6-12
| |
| | style="text-align:center" | [[Aβ]]
| |
| | style="text-align:center" | 36-72
| |
| |-
| |
| | style="text-align:center" | III
| |
| | style="text-align:center" | 1-6
| |
| | style="text-align:center" | [[Aδ]]
| |
| | style="text-align:center" | 4-36
| |
| |-
| |
| | style="text-align:center" | -
| |
| | style="text-align:center" | ≦3
| |
| | style="text-align:center" | [[B(注1)]]
| |
| | style="text-align:center" | 3-15
| |
| |-
| |
| | style="text-align:center" | 無髄神経
| |
| | style="text-align:center" | IV
| |
| | style="text-align:center" | 0.2-1.5
| |
| | style="text-align:center" | [[C]]
| |
| | style="text-align:center" | 0.4-2.0
| |
| |}
| |
| (注1)B繊維は一般的に自律神経系前繊維を指す。
| |
|
| |
|
| {| cellspacing="1" cellpadding="1" border="1"
| | [[Image:Node of Ranvier.png|RTENOTITLE]]<br> |
| |+'''表2.神経線維の受容器の分類とその様式'''<ref name="ref1" />
| |
| |-
| |
| | style="text-align:center" | 受容器の種類
| |
| | style="text-align:center" | 伝達速度からの分類
| |
| | style="text-align:center" | 太さあるいは機能からの名称
| |
| | style="text-align:center" | 様式
| |
| |-
| |
| | 皮膚と皮下の[[機械受容器]]
| |
| |
| |
| |
| |
| | [[触覚]]
| |
| |-
| |
| | [[マイスナー小体]]
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | RA1 (注2)
| |
| | なでる、粗動
| |
| |-
| |
| | [[メルケル細胞]]
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | SA1 (注3)
| |
| | 圧力、触感
| |
| |-
| |
| | [[パチニ小体]]
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | RA2 (注2)
| |
| | 振動
| |
| |-
| |
| | [[ルフィニ終末]]
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | SA2 (注3)
| |
| | 皮膚の伸縮
| |
| |-
| |
| | [[Hair-tylotrich]], [[hair-guard]]
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | G1、G2
| |
| | なでる、粗動
| |
| |-
| |
| | [[Hair-down]]
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | D
| |
| | 軽くなでる
| |
| |-
| |
| | Field
| |
| | style="text-align:center" | Aα、β
| |
| | style="text-align:center" | F
| |
| | 皮膚の伸縮
| |
| |-
| |
| | C機械受容器
| |
| | style="text-align:center" | C
| |
| |
| |
| | なでる、erotic touch
| |
| |-
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| |-
| |
| | [[温度受容器]]
| |
| |
| |
| |
| |
| | 温度
| |
| |-
| |
| | [[冷覚受容器]]
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | III
| |
| | 皮膚冷感(<25℃[77℉])
| |
| |-
| |
| | [[温覚]]受容器
| |
| | style="text-align:center" | C
| |
| | style="text-align:center" | IV
| |
| | 皮膚温感(>35℃[95℉])
| |
| |-
| |
| | [[高熱侵害受容器]]
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | III
| |
| | 高温(>45℃[113℉])
| |
| |-
| |
| | [[寒冷侵害受容器]]
| |
| | style="text-align:center" | C
| |
| | style="text-align:center" | IV
| |
| | 低温(<5℃[41℉])
| |
| |-
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| |-
| |
| | [[侵害受容器]]
| |
| |
| |
| |
| |
| | 痛み
| |
| |-
| |
| | 機械的
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | III
| |
| | 鋭く穿刺するような痛み
| |
| |-
| |
| | 温度機械的(熱)
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | III
| |
| | 焼けるような痛み
| |
| |-
| |
| | 温度機械的(寒冷)
| |
| | style="text-align:center" | C
| |
| | style="text-align:center" | IV
| |
| | 凍てつく痛み
| |
| |-
| |
| | 多様式的
| |
| | style="text-align:center" | C
| |
| | style="text-align:center" | IV
| |
| | 鈍い焼けるような痛み
| |
| |-
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| | style="background-color:#a9a9a9" |
| |
| |-
| |
| | 筋・骨格の機械受容器
| |
| |
| |
| |
| |
| | 四肢の[[固有受容性感覚]]
| |
| |-
| |
| | [[筋紡錘]](第一)
| |
| | style="text-align:center" | Aα
| |
| | style="text-align:center" | Ia
| |
| | 筋肉の長さとスピード
| |
| |-
| |
| | 筋紡錘(第二)
| |
| | style="text-align:center" | Aβ
| |
| | style="text-align:center" | II
| |
| | 筋肉の伸展
| |
| |-
| |
| | [[ゴルジ腱器官]]
| |
| | style="text-align:center" | Aα
| |
| | style="text-align:center" | Ib
| |
| | 筋肉の収縮
| |
| |-
| |
| | [[間接嚢受容器]]
| |
| | style="text-align:center" | Aβ
| |
| | style="text-align:center" | II
| |
| | 筋肉の角度
| |
| |-
| |
| | [[伸縮感応性自由終末]]
| |
| | style="text-align:center" | Aδ
| |
| | style="text-align:center" | III
| |
| | 過剰な伸展あるいは力
| |
| |}
| |
|
| |
|
| (注2)Rapidly adapting type 1 あるいは 2
| | 図2:有髄神経の構造とサブドメイン |
|
| |
|
| (注3)Slowly adapting type1 あるいは 2
| | <br> |
|
| |
|
| == 有髄線維の構造 ==
| |
|
| |
|
| [[Image:Node of Ranvier.png|thumb|350px|<b>図1.有髄線維の構造とサブドメイン</b>]]
| | == 3:跳躍伝導(saltatory conduction) == |
| | <br> |
|
| |
|
| 髄鞘は細胞[[形質膜]]の多層構造体であるため、[[細胞形質膜]]や細胞内小胞膜などの他の多くの細胞と比べてタンパク質成分が少なく、脂質が約8割程度を占め、残りがタンパク質である。髄鞘を構成する主な脂質は糖脂質[[ガラクトセレブロシド]]([[Galactocerebroside]])とその硫酸化誘導体[[スルファチド]]([[Sulfatide]])である<ref><pubmed>9530920</pubmed></ref>。神経軸索の髄鞘と髄鞘の隙間は特別な名称がつけられており、非常に特異な構造をしている。[[ノード]](node)に[[電位作動型ナトリウムチャネル]][[ナトリウムチャネル#.CE.B1.E3.82.B5.E3.83.96.E3.83.A6.E3.83.8B.E3.83.83.E3.83.88|Nav1.2]]や[[ナトリウムチャネル#.CE.B1.E3.82.B5.E3.83.96.E3.83.A6.E3.83.8B.E3.83.83.E3.83.88|Nav1.6]]、[[アンキリン]]G(Ankyrin G)などが、[[パラノード]]([[paranode]])には[[Caspr]]など、[[ジャクスタパラノード]]([[jaxtaparanode]])には[[カリウムチャネル#遅延整流性カリウム電流|Kv1.2]]などの[[カリウムチャネル]]などが分布する<ref><pubmed>18803321</pubmed></ref>。発達とともにこれらのチャネルやそのアイソフォームの分布が変化する<ref><pubmed>9880595</pubmed></ref>。また、病態時にもチャネルの分布は変化する<ref><pubmed>12898531</pubmed></ref> <ref><pubmed>10739574</pubmed></ref>。
| | 神経軸索の起始部で髄鞘に覆われていない部分は初節(axon initial segment; AISと略される)とよばれ、電位依存性のナトリウムチャネルが高密度に集中しており、活動電位が最初に発火する部分である。AIS以降の神経軸索の遠位部では、ほぼ等間隔に髄鞘が途切れる節があり、その部分をランヴィエの絞輪とよぶ。髄鞘間のランヴィエの絞輪には電位依存性ナトリウムチャネルが存在し、ここで活動電位は再生される。ある節に起こった脱分極は、受動的伝播によって即座に次の節に伝わり、活動電位は髄鞘化された神経軸索上を節から節へ伝わっていくので、跳躍伝導と呼ばれる。この伝播様式は活動電位が速く伝わるうえ、興奮が軸索の細胞膜上の狭いランヴィエの絞輪に限定されるので、代謝エネルギーの節約にもなる。 |
|
| |
|
| 中枢神経系(オリゴデンドロサイト)と末梢神経系(シュワン細胞)で髄鞘の巻き方が異なる。オリゴデンドロサイトは離れた複数の軸索個々に髄鞘を形成するのに対し、シュワン細胞はいくつかの軸索を抱え込むようにして包んだ後、1本の軸索を選別して、その周りに髄鞘を形成する<ref><pubmed>22192173</pubmed></ref>。髄鞘を構成するタンパク質も異なる。中枢神経系では髄鞘の主要構成タンパク質はPLP,MBPである。一方、末梢神経系では[[P0]](ピーゼロ)と[[P2]]が主要構成タンパク質である。その他のタンパク質は中枢神経系ではCNPase、MOG、[[Mag|MAG]]などがあり、末梢神経系では[[PMP22]]、MAG、MOGなどが発現している。
| | [[Image:跳躍伝導.png|図2:興奮伝達(活動電位の伝達)]] [[Image:跳躍伝導(有髄).png|400x220px]]<br> |
|
| |
|
| [[Image:受動輸送と能動輸送(改).jpg|thumb|350px|'''図2.興奮伝達(活動電位の伝達)<ref>ニューロンの生理学 D. Tritsch, D.Chenoy-Marchais, A.Feltz</ref>''']]
| | 図3:興奮の伝達 (活動電位の伝達) 図4:跳躍伝導 (髄鞘が絶縁体の役割を果たし、活動電位の伝達が)<br> <br> |
| == 跳躍伝導==
| |
| [[Image:跳躍伝導(有髄).png|thumb|350px|'''図3.跳躍伝導'''<br>髄鞘が絶縁体の役割を果たし、活動電位の伝達が早くなる。]]
| |
|
| |
|
| それぞれのイオンが受動的に流失し、[[電気化学的平衡]]が失われた場合、それを補償する[[能動的輸送]]が必要となる。この能動的輸送にはエネルギーが必要とされる。一般的には一次と二次[[能動輸送]]という二種類の重要な能動輸送がある(図2)。
| | <br> |
| 神経軸索の起始部で髄鞘に覆われていない部分は[[初節]](axon initial segment; AISと略される)とよばれ、電位依存性ナトリウムチャネルが高密度に集積しており、通常[[活動電位]]が最初に発火する<ref><pubmed>20631711</pubmed></ref>。AISで発現するチャネルは発達に伴い、あるいは病態時にその発現や分布が変化する<ref><pubmed>22103418</pubmed></ref>。AIS以降の軸索では、ほぼ等間隔にランヴィエ絞輪が現れる。AISやランヴィエ絞輪で発生した活動電位は、受動的伝播(電気緊張電位)によって次のランヴィエ絞輪に伝わる。髄鞘の絶縁性が高いために電位変化の減衰度合いは小さく押さえられる。そのため無髄軸索よりも離れた距離でも臨界脱分極を越える。またランヴィエ絞輪では電位依存性ナトリウムチャネルが集積しているため、活動電位が発生しやすい。この2つの効果によって跳躍伝導(saltatory conduction)が起こる。跳躍伝導は、活動電位の伝導速度を速く保つとともに、活動電位発生がランヴィエ絞輪に限局されるために、代謝エネルギーの節約にもなる。
| |
|
| |
|
| == 脱髄性疾患== | | == 4:脱髄性疾患(Demyelinating disease) == |
|
| |
|
| 正常な発生における髄鞘形成がなされたのち、神経軸索から髄鞘が脱落することを脱髄という<ref><pubmed>18558866</pubmed></ref>。[[脱髄性疾患]](demyelinating disease)では、髄鞘の消失により神経伝導速度が遅くなり、さまざまな神経症状が引き起こされる。脱髄が起こる場所により症状はさまざまである。[[wj:運動麻痺|運動麻痺]]、[[wj:感覚麻痺|感覚麻痺]]、[[wj:視力障害|視力障害]]などが起こる。
| | 正常な発生における髄鞘形成がなされたのち、神経軸索から髄鞘が脱落すること[よしむら8]。この疾患を脱髄性疾患(demyelinating disease)といい、髄鞘の消失により神経伝導速度が遅くなり、さまざまな神経症状が引き起こされる。脱髄が起こる場所により症状はさまざまである。運動麻痺、感覚麻痺、視力障害などが起こる。中枢神経系では多発性硬化症(Multiple sclerosis)が、末梢神経系ではギラン・バレー症候群(Guillain-Barré syndrome)などがある。また、髄鞘形成が不完全な髄鞘形成不全疾患(dysmyelination disease)とは区別される。 <br> |
|
| |
|
| 中枢性脱髄疾患の中では[[多発性硬化症]](multiple sclerosis, MS)が最も多くみられる。日本での有病率は2006年では10万人あたり8 - 9人程度と推定されている。若年成人での神経障害の主な原因である。約80-90%のMS患者が二十代後半で再発-[[寛解]]型になり、寛解期には共通して、ある一定程度神経障害が一過的に回復する。発病から数年後、多くの患者が実質上機能回復はなくなり第二ステージに移り、病状は進行型になる。そして、あとの残り10-20%の患者は初めから寛解を経験することのない進行型である<ref name=ref10><pubmed>22189514</pubmed></ref>。[[カルシウムチャネル]]ブロッカーである[[Fampridine]](Fampyra)はMS治療において初めて機能回復に成功した経口投与薬であるが、転倒する、背中が痛む、[[めまい]]がする、[[不眠]]、[[疲労感]]、[[悪心]]、[[平衡障害]]などの副作用を伴うことが知られている<ref><pubmed>7354839</pubmed></ref> <ref><pubmed>6631441</pubmed></ref> <ref><pubmed>3001584</pubmed></ref> <ref><pubmed>2435223</pubmed></ref>。[[wj:インターフェロン|インターフェロン]]β(Interferon β)と[[wj:グラチマラー酢酸塩|グラチマラー酢酸塩]](glatiramer acetate)が現在もっともよく用いられている。これらの薬剤は根本治療法であるが、おもに再発-寛解型の患者に用いられている<ref name=ref10 />。
| | <br> |
|
| |
|
| また、末梢神経系では[[ギラン・バレー症候群]]([[Guillain-Barré syndrome]])や[[慢性炎症性脱髄性多発神経炎]]([[chronic inflammatory demyelinating polyneuropathy]]; [[CIDP]])、などがある。また、最近の研究により[[統合失調症]]との関連が示唆されている<ref><pubmed>18538868</pubmed></ref> <ref><pubmed>20216548</pubmed></ref> <ref><pubmed>21196354</pubmed></ref>。髄鞘形成が不完全な[[髄鞘形成不全疾患]](dysmyelinating disease)とは区別される。
| | ~大きく分けて中枢神経系と末梢神経系の疾患がある~ |
|
| |
|
| {| width="583" cellspacing="1" cellpadding="1" border="1"
| | ○中枢神経系<br> ・多発性硬化症<br> ・視神経脊髄炎(Devic症候群)<br> ・同心円硬化症(Balo病)<br> ・急性散在性脳脊髄炎(ADEM)<br> ・炎症性広汎性硬化症(Schilder病)<br> ・感染性<br> ・亜急性硬化症全脳炎(SSPE)<br> ・進行性多巣性白質脳症(PML)<br> ・中毒・代謝性<br> ・低酸素脳症<br> ・橋中心髄鞘破壊症<br> ・ビタミンB12欠乏症<br> ・血管性<br> ・Binswanger病<br> ○末梢神経系<br> ・ギラン・バレー症候群<br> ・フィッシャー症候群<br> ・慢性炎症性脱髄性多発根神経炎<br><br> <br> |
| |-
| |
| | rowspan="9" | 中枢神経系
| |
| | rowspan="3" | 原発性
| |
| | [[多発性硬化症]]
| |
| | [[同心円硬化症]]([[Balo病]])
| |
| |-
| |
| | [[急性散在性脳脊髄炎]]([[ADEM]])
| |
| |-
| |
| | [[炎症性広汎性硬化症]]([[Schilder病]])
| |
| |-
| |
| | rowspan="2" | 感染性
| |
| | [[亜急性硬化症全脳炎]](SSPE)
| |
| |-
| |
| | [[進行性多巣性白質脳症]](PML)
| |
| |-
| |
| | rowspan="3" | 中毒・代謝性
| |
| | [[低酸素脳症]]
| |
| |-
| |
| | [[橋中心髄鞘破壊症]]
| |
| |-
| |
| | [[wj:ビタミンB12|ビタミンB12]]欠乏症
| |
| |-
| |
| | 血管性
| |
| | [[Binswanger病]]
| |
| |-
| |
| | rowspan="2" | 末梢神経系
| |
| | [[ギラン・バレー症候群]]
| |
| | [[フィッシャー症候群]]
| |
| |-
| |
| | colspan="2" | [[慢性炎症性脱髄性多発神経炎]]
| |
| |}
| |
| | |
| '''表3.代表的な脱髄性疾患'''
| |
| | |
| 大きく分けて中枢神経系と末梢神経系の疾患がある。
| |
|
| |
|
| == 関連語 == | | == 関連語 == |
|
| |
|
| *[[グリア細胞]] | | *[[グリア細胞]] |
| | |
| *[[オリゴデンドロサイト]] | | *[[オリゴデンドロサイト]] |
| | |
| *[[シュワン細胞]] | | *[[シュワン細胞]] |
| | |
| *[[活動電位]] | | *[[活動電位]] |
| | |
| *[[軸索]] | | *[[軸索]] |
| *[[跳躍伝導]]
| | |
| *[[ランビエ絞輪]] | | *[[ランビエ絞輪]] |
| *[[脱髄性疾患]]
| |
|
| |
|
| == 参考文献 == | | == 参考文献 == |
| | 1:Jorge A Pereira et al., Trends Neurosci.:35(2); 123-34 (2012) Molecular mechanisms regulating myelination in the peripheral nervous system. [http://www.ncbi.nlm.nih.gov/pubmed?term=pereira%20molecular%202012%20mechanisms%20myelination]<Br> |
| | 2:James L. Salzer et al., Glia 56:1532-1540 (2008) Molecular Domains of Myelinated Axons in the Peripheral Nervous System [http://www.ncbi.nlm.nih.gov/pubmed/18803321]<br> |
| | |
| | 3:Shelly A. Buffington et al., European Journal of Neuroscience, Vol. 34, pp. 1609-1619 (2011) The axon initial segment in nervous system disease and injury. [http://www.ncbi.nlm.nih.gov/pubmed/22103418]<br> |
|
| |
|
| <references />
| | (執筆者:清水崇弘、池中一裕 担当編集委員:柚崎通介) |
英語1: Myelinated nerve 英語2: Medullated nerve
中枢神経系では希突起膠細胞(あるいはオリゴデンドロサイト[oligodendrocyte])、末梢神経系ではシュワン細胞(Schwann cell)というグリア細胞の形成する髄鞘(ミエリン)[参照:「髄鞘」[1]]が神経軸索[参照:「軸索」[[2]]]の周囲に形成された神経のことをいう。神経軸索は樹状突起と比べると細く、長く、平滑な細胞突起である。ミエリン鞘は等間隔にあるランヴィエの絞輪(node of Ranvier)で途切れていて、軸索中のNa+チャネルはほぼこの節に集中している。
髄鞘はリン脂質に富んだタンパク質で層構造を形成しているため、髄鞘は絶縁体の役割を果たし、軸索膜を絶縁して膜からの電流のもれをほぼ完全に防ぎ、神経細胞からの電気信号を跳躍伝導させることができる。また、単に絶縁体としての働きだけではなく、神経軸索の保護や軸索との間に緊密な相互作用を行うことで、さまざまな神経機能を調節している。主要構成蛋白質はミエリン塩基性蛋白質(Myelin basic protein; MBP)、プロテオリピドプロテイン(myelin proteolipid protein; PLP)ミエリン関連糖タンパク質(Myelin associated glycoprotein; MAG)、ミエリンオリゴデンドロサイト糖タンパク質(Myelin oligodendrocyte glycoprotein; MOG)などがある。
1:神経の分類と髄鞘の有無
末梢神経の神経線維は髄鞘の有無、直径、伝導速度等で分類される。有髄線維と無髄線維では有髄線維が、同じ種類の線維間では直径が大きい方が伝導速度が速い。前者は跳躍伝導により、後者は電気緊張電位の広がりの違いにより速い速度を得ている。一般に骨格筋運動と付随する固有感覚、部位のはっきりした皮膚感覚は伝導速度の速い線維を、交感神経活動や鈍痛などは伝導速度の遅い線維を利用して伝えられる。
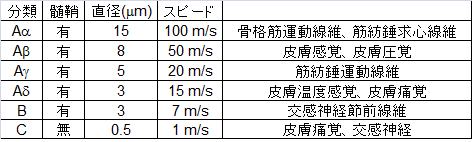
図1:神経繊維の分類と髄鞘の有無
2:有髄神経の構造
神経軸索の髄鞘と髄鞘の隙間は特別な名称がつけられており、非常に特異な構造をしている。ノード(node)に電位作動型イオンチャネルNav1.2や Nav1.6、アンキリンG(AnkG)などが、パラノード(paranode)にはCasprなど、ジャクスタパラノード(jaxtaparanode)にはKv1.2などのカリウムチャネルなどが分布する。中枢神経系(オリゴデンドロサイト)と末梢神経系(シュワン細胞)で髄鞘の巻き方が少し異なる。オリゴデンドロサイトは離れた軸索に別々に髄鞘を形成するのに対し、シュワン細胞はいくつかの軸索を抱え込むようにして包んだ後、1本の軸索を選別して、その周りに髄鞘を形成する。[]髄鞘を構成する蛋白質も少し異なる。
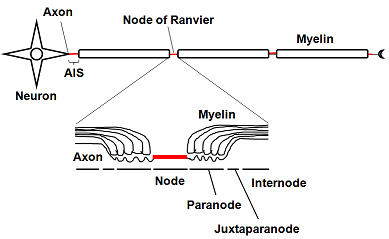
図2:有髄神経の構造とサブドメイン
3:跳躍伝導(saltatory conduction)
神経軸索の起始部で髄鞘に覆われていない部分は初節(axon initial segment; AISと略される)とよばれ、電位依存性のナトリウムチャネルが高密度に集中しており、活動電位が最初に発火する部分である。AIS以降の神経軸索の遠位部では、ほぼ等間隔に髄鞘が途切れる節があり、その部分をランヴィエの絞輪とよぶ。髄鞘間のランヴィエの絞輪には電位依存性ナトリウムチャネルが存在し、ここで活動電位は再生される。ある節に起こった脱分極は、受動的伝播によって即座に次の節に伝わり、活動電位は髄鞘化された神経軸索上を節から節へ伝わっていくので、跳躍伝導と呼ばれる。この伝播様式は活動電位が速く伝わるうえ、興奮が軸索の細胞膜上の狭いランヴィエの絞輪に限定されるので、代謝エネルギーの節約にもなる。
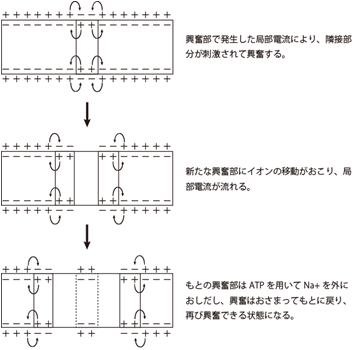
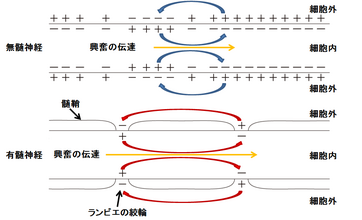
図3:興奮の伝達 (活動電位の伝達) 図4:跳躍伝導 (髄鞘が絶縁体の役割を果たし、活動電位の伝達が)
4:脱髄性疾患(Demyelinating disease)
正常な発生における髄鞘形成がなされたのち、神経軸索から髄鞘が脱落すること[よしむら8]。この疾患を脱髄性疾患(demyelinating disease)といい、髄鞘の消失により神経伝導速度が遅くなり、さまざまな神経症状が引き起こされる。脱髄が起こる場所により症状はさまざまである。運動麻痺、感覚麻痺、視力障害などが起こる。中枢神経系では多発性硬化症(Multiple sclerosis)が、末梢神経系ではギラン・バレー症候群(Guillain-Barré syndrome)などがある。また、髄鞘形成が不完全な髄鞘形成不全疾患(dysmyelination disease)とは区別される。
~大きく分けて中枢神経系と末梢神経系の疾患がある~
○中枢神経系
・多発性硬化症
・視神経脊髄炎(Devic症候群)
・同心円硬化症(Balo病)
・急性散在性脳脊髄炎(ADEM)
・炎症性広汎性硬化症(Schilder病)
・感染性
・亜急性硬化症全脳炎(SSPE)
・進行性多巣性白質脳症(PML)
・中毒・代謝性
・低酸素脳症
・橋中心髄鞘破壊症
・ビタミンB12欠乏症
・血管性
・Binswanger病
○末梢神経系
・ギラン・バレー症候群
・フィッシャー症候群
・慢性炎症性脱髄性多発根神経炎
関連語
参考文献
1:Jorge A Pereira et al., Trends Neurosci.:35(2); 123-34 (2012) Molecular mechanisms regulating myelination in the peripheral nervous system. [3]
2:James L. Salzer et al., Glia 56:1532-1540 (2008) Molecular Domains of Myelinated Axons in the Peripheral Nervous System [4]
3:Shelly A. Buffington et al., European Journal of Neuroscience, Vol. 34, pp. 1609-1619 (2011) The axon initial segment in nervous system disease and injury. [5]
(執筆者:清水崇弘、池中一裕 担当編集委員:柚崎通介)



.png)