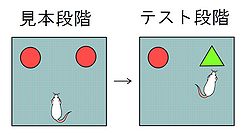
Ennaceur & Delacour (1988)<ref name="Enna" /> は45 cm x 65cmで高さ45 cmの壁のある実験アリーナに、2つの同じ物体を置き、これを[[wikipedia:ja:ラット|ラット]]に数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度、動物を2つの物体のある広場に戻し探索させた([[テスト]]段階)。一方は遅延前に提示した物体と同じ物体(馴染物体)で、他方は異なる物体(新奇物体)である(図1)。新奇物体の探索時間が馴染物体の探索時間より長ければ、動物が以前に探索した物体を認知したと結論できる。
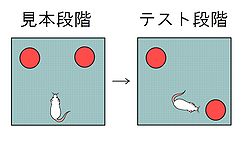
Dix & Aggleton(1999)<ref name="Dix" />は100 cm x 100cmで高さ46 cmの壁のある実験アリーナに、2つの同じ物体を置き、これをラットに数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度動物を2つの物体のある広場に戻し探索させた(テスト段階)。この時、使用する2つの物体は見本段階と同じものであるが、片方の物体のみ、見本とは異なる位置に配置した(図2)。もし新しい位置に移動した物体への探索行動が増加すれば、物体の位置関係についての認知的処理が行われたとみなすことができる。
探索行動の変化は物体の配置の変化の仕方によって異なる<ref name="Thi1987">'''Thinus-Blanc, C., L. Bouzouba, K. Chaix, N. Chapuis, M. Durup, & B. Poucet'''<br>A study of spatial parameters encoded during exploration in hamsters''<br>''Journal of Experimental Psychology: [[Animal]] Behavior Processes'':1987,13,418-427</ref>。例えば、4つの物体を配置して馴致した後、第3セッションで1つの物体の配置を変化させると、配置が変わった物体に対して探索時間が増加した。
動物に二つの同じ物体を探索させた後、ひとつの物体を新しい物体に置き換えると、正常な動物は新奇物体を優先して長時間探索する<ref name="Enna" /><ref name="Dix"><pubmed>10512585</pubmed></ref> 。Ennaceur & Delacour (1988)<ref name="Enna" /> は45 cm x 65cmで高さ45 cmの壁のある実験アリーナに、2つの同じ物体を置き、これをラットに数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度、動物を2つの物体のある広場に戻し探索させた(テスト段階)。一方は遅延前に提示した物体と同じ物体(馴染物体)で、他方は異なる物体(新奇物体)である(図1)。新奇物体の探索時間が馴染物体の探索時間より長ければ、動物が以前に探索した物体を認知したと結論できる。脳損傷や薬物投与によって馴染物体と新奇物体の探索時間に違いが見られなくなった場合、物体認知の障害が生じていると解釈できる。また、遅延時間に依存して障害が生じる場合には、作業記憶障害が生じている可能性を検討すべきである。この課題は課題のルールに関する学習が必要でないため、参照記憶障害の可能性は除外できるだろう。
==== 環境の変化 ====
==== 位置の変化[[Image:Uekita Ob Fig.2.jpg|right|250px|図2 位置(空間)認知を調べる課題]] ====
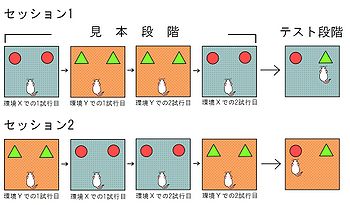
[[Image:Uekita Ob Fig.4 new.jpg|thumb|350px|'''図4.環境の認知を調べる課題'''<br>(<ref name="Dix" />をもとに作成)丸は物体A、三角は物体Bを示す。]]
あらかじめ探索させた複数の物体の配置を変化させると、正常な動物では配置の変化した物体に対して探索行動が増加する<ref name="Dix" />。Dix & Aggleton(1999)<ref name="Dix" />は100 cm x 100cmで高さ46 cmの壁のある実験アリーナに、2つの同じ物体を置き、これをラットに数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度動物を2つの物体のある広場に戻し探索させた(テスト段階)。この時、使用する2つの物体は見本段階と同じものであるが、片方の物体のみ、見本とは異なる位置に配置した(図2)。もし新しい位置に移動した物体への探索行動が増加すれば、物体の位置関係についての認知的処理が行われたとみなすことができる。
探索行動の変化は物体の配置の変化の仕方によって異なる<ref name="Thi1987">'''Thinus-Blanc, C., L. Bouzouba, K. Chaix, N. Chapuis, M. Durup, & B. Poucet'''<br>A study of spatial parameters encoded during exploration in hamsters''<br>''Journal of Experimental Psychology: Animal Behavior Processes'':1987,13,418-427</ref>。例えば、4つの物体を配置して馴致した後、第3セッションで1つの物体の配置を変化させると、配置が変わった物体に対して探索時間が増加した。4つの物体の配置を4角形から3角形へと幾何学的に変化させると、配置が変わった物体と変わっていない物体いずれに対しても探索量が増加した。興味深いことに、幾何学的配置を保ったまま、物体間の距離のみが変わった場合には探索時間の増加はみられなかった。また、1つの物体を取り除くと、残った物体への探索量が増加した。
物体の位置関係の認知と物体そのものの認知(2.2.1)やこれらに関わる神経基盤は、物体馴致セッション、空間認識テスト、物体認[[Image:Uekita Ob Fig.3.jpg|right|400px|図3 場所と物体の複合課題 (Save et al. (1992)をもとに作成)]]識テストを含む一連の手続きによって同時に検討することができる(図3)。Save, Poucet, Foreman, & Buhot (1992)<ref><pubmed>1616611</pubmed></ref> は、円形の実験アリーナに5つの異なる物体を配置し、6分間これをラットに探索させた。全ての物体に対して馴染みを形成するため、物体馴致を3セッション繰り返した後、空間認識テストにおいて2個の物体を移動させた。統制群と前部頭頂皮質損傷群のラットは配置の変化した物体に対して変化していない物体よりも多く探索行動を示したが、海馬損傷群と後部頭頂皮質損傷群のラットではこのような傾向が見られず、物体の位置関係の認知に失敗した。次の物体認識テストでは、一つの物体を新しい物体に置き換えたところ、全ての群のラットが新しい物体に対して探索行動が増加した。これらの結果は、海馬や後部頭頂皮質が物体の位置関係の認知に関与するが、物体自体の認知には関与しない事を示している。位置関係の認知に選択的な障害は、げっ歯類デグーの海馬破壊<ref><pubmed>21291914</pubmed></ref>やラットNMDA受容体の薬理学的阻害<ref>'''関口理久子'''<br>ラットの空間探索行動に及ぼすNMDAアンタゴニスト,MK-801の効果''<br>''心理学研究'':1997,68,88-94</ref>によっても生じることが報告されている。
新奇物体への嗜好性は、単純に新奇物体と馴染物体の探索量を比較することでも可能であるが、探索量の個体差の問題を除外するために、馴染物体と新奇物体の全探索量に対する新奇物体探索量の割合として示されることが多い。物体馴致セッションにおける探索量とテストにおける馴染物体または新奇物体の探索量の変化量を指標とすることもある。統計的に有意な嗜好性を示しているかどうかについて、ワンサンプルt検定により、群の平均探索率がチャンスレベルよりも有意に異なっているかどうかを調べることができる。ただし、新奇物体への嗜好性が強いということが、認知能力の高さを示しているかどうかは明らかでない<ref name="Whi" />。また、前述のThinus-Blanc et al.(1987)<ref name="Thi1987" />の実験によると、物体の配置の変化の仕方によって、配置が変わった物体だけでなく、配置が変わっていない物体に対しても探索量が増えることがある。したがって、位置関係の認知的処理ができているかどうかの判断は、単純に移動物体と固定物体の比較だけでは不十分であるだろう。
また、前述のThinus-Blanc et al.(1987)<ref name="Thi1987" />の実験によると、物体の配置の変化の仕方によって、配置が変わった物体だけでなく、配置が変わっていない物体に対しても探索量が増えることがある。したがって、位置関係の認知的処理ができているかどうかの判断は、単純に移動物体と固定物体の比較だけでは不十分であるだろう。
== 物体認知の神経基盤 ==
==ヒトにおける物体認知の発達==
上述のように、物体探索課題を用いた海馬損傷実験では、海馬は物体の配置や物体の置かれた環境の符号化に必要であるが、物体認知そのものには重要でないと考えられている。これまでのところ、物体自体の認知に関わる脳領域について、物体探索課題では積極的な結果が得られていない。しかし、物体認知記憶を測定する遅延非見本合わせ課題(delayed nonmatching to sample,DNMS)を用いた損傷研究により、嗅皮質(rhinal cortex)が物体認知に重要であると考えられている。この課題は、ある刺激を前に見たかどうかについての物体認知記憶を測定するために使用される。見本試行において、テーブル上に見本物体が短時間提示され、遅延時間後の選択試行では、見本物体と同じ物体が新奇物体とともに提示される。動物が新奇物体を選択すると報酬が与えられる。
これまで、物体探索行動を指標として測定された物体認知の神経基盤としては、海馬が物体の空間的な位置情報処理に関与するが、物体認知そのものには重要でないという一致した見解が得られてきた。主にげっ歯類を対象とする物体探索課題では、物体認知に関わる脳領域についての積極的な結果が得られていない。霊長類では非見本合わせ(nonmatching to sample)という手続きにより物体認知が測定されてきた。この課題の物体認知テストでは、見本試行において、テーブル上に見本物体が短時間提示され、選択試行では見本物体と同じ物体が新奇物体とともに提示される。動物が新奇物体を選択すると報酬が与えられる。この課題の位置認知テストでは、テーブル上の左右におかれたふた付の報酬ケースのうち、ランドマークとなる物体により近い方を選択すると報酬が得られる。下側頭葉 TE 野を損傷されたサルは、物体認知テストにおいて著しい障害を示したが、位置認知テストでは障害を示さなかった。一方、下頭頂酔葉PG野を損傷されたサルは、位置認知テストで障害を示したが、物体認知テストでは障害を示さなかった。このような結果から、Mishkinらは、同一の物体の情報処理が、処理内容によって異なる経路を介することを主張した<ref><pubmed>7126325</pubmed></ref> 。すなわち、視覚的に提示された物体について、空間的位置情報は一次視覚野から背側に流れる経路「where pathway」で処理され、物体認知は一次視覚野から腹側に流れる経路「what pathway」で処理されるとした。
また側頭葉内側部と物体記憶の関連について、見本試行と選択試行の間に遅延時間を設定する[[遅延非見本合わせ課題]](delayed nonmatching to sample)によって検討されてきた。この課題を用いた初期の実験<ref><pubmed>418358</pubmed></ref>では、[[wikipedia:ja:マカクザル|マカクザル]]の海馬と[[扁桃体]]を含む側頭葉内側部の損傷の効果が検討された。見本試行と選択試行の遅延時間が10秒以内である場合、この課題の遂行に損傷の影響はなかったが、それよりも長い遅延時間が挿入されると、その時間依存的に課題の正答率が低くなった。後に同研究者によって損傷の精度を高めて追試が行われた結果、この障害は海馬や扁桃体単独の損傷では生じず、むしろそれらの近辺領域にある嗅皮質の損傷が障害を引き起こしたことが明らかになった<ref><pubmed>9698344</pubmed></ref>。したがって、物体認知記憶には海馬や扁桃体ではなく[[嗅皮質]]が関与すると考えられる。
動物に二つの同じ物体を探索させた後、ひとつの物体を新しい物体に置き換えると、正常な動物は新奇物体を優先して長時間探索する[5][7] 。Ennaceur & Delacour (1988)[5] は45 cm x 65cmで高さ45 cmの壁のある実験アリーナに、2つの同じ物体を置き、これをラットに数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度、動物を2つの物体のある広場に戻し探索させた(テスト段階)。一方は遅延前に提示した物体と同じ物体(馴染物体)で、他方は異なる物体(新奇物体)である(図1)。新奇物体の探索時間が馴染物体の探索時間より長ければ、動物が以前に探索した物体を認知したと結論できる。脳損傷や薬物投与によって馴染物体と新奇物体の探索時間に違いが見られなくなった場合、物体認知の障害が生じていると解釈できる。また、遅延時間に依存して障害が生じる場合には、作業記憶障害が生じている可能性を検討すべきである。この課題は課題のルールに関する学習が必要でないため、参照記憶障害の可能性は除外できるだろう。
位置の変化図2 位置(空間)認知を調べる課題
あらかじめ探索させた複数の物体の配置を変化させると、正常な動物では配置の変化した物体に対して探索行動が増加する[7]。Dix & Aggleton(1999)[7]は100 cm x 100cmで高さ46 cmの壁のある実験アリーナに、2つの同じ物体を置き、これをラットに数分間探索させた(見本段階)。一旦、ラットを広場から出して遅延をおき、再度動物を2つの物体のある広場に戻し探索させた(テスト段階)。この時、使用する2つの物体は見本段階と同じものであるが、片方の物体のみ、見本とは異なる位置に配置した(図2)。もし新しい位置に移動した物体への探索行動が増加すれば、物体の位置関係についての認知的処理が行われたとみなすことができる。
新奇物体への嗜好性は、単純に新奇物体と馴染物体の探索量を比較することでも可能であるが、探索量の個体差の問題を除外するために、馴染物体と新奇物体の全探索量に対する新奇物体探索量の割合として示されることが多い。物体馴致セッションにおける探索量とテストにおける馴染物体または新奇物体の探索量の変化量を指標とすることもある。統計的に有意な嗜好性を示しているかどうかについて、ワンサンプルt検定により、群の平均探索率がチャンスレベルよりも有意に異なっているかどうかを調べることができる。ただし、新奇物体への嗜好性が強いということが、認知能力の高さを示しているかどうかは明らかでない[6]。また、前述のThinus-Blanc et al.(1987)[8]の実験によると、物体の配置の変化の仕方によって、配置が変わった物体だけでなく、配置が変わっていない物体に対しても探索量が増えることがある。したがって、位置関係の認知的処理ができているかどうかの判断は、単純に移動物体と固定物体の比較だけでは不十分であるだろう。
物体認知の神経基盤
上述のように、物体探索課題を用いた海馬損傷実験では、海馬は物体の配置や物体の置かれた環境の符号化に必要であるが、物体認知そのものには重要でないと考えられている。これまでのところ、物体自体の認知に関わる脳領域について、物体探索課題では積極的な結果が得られていない。しかし、物体認知記憶を測定する遅延非見本合わせ課題(delayed nonmatching to sample,DNMS)を用いた損傷研究により、嗅皮質(rhinal cortex)が物体認知に重要であると考えられている。この課題は、ある刺激を前に見たかどうかについての物体認知記憶を測定するために使用される。見本試行において、テーブル上に見本物体が短時間提示され、遅延時間後の選択試行では、見本物体と同じ物体が新奇物体とともに提示される。動物が新奇物体を選択すると報酬が与えられる。
↑Ennaceur, A. (2010).
One-trial object recognition in rats and mice: methodological and theoretical issues. Behavioural brain research, 215(2), 244-54.
[PubMed:20060020]
[WorldCat]
[DOI]
↑Berlyne, D.E. (1966).
Curiosity and exploration. Science (New York, N.Y.), 153(3731), 25-33.
[PubMed:5328120]
[WorldCat]
[DOI]
↑Poucet, B. (1989).
Object exploration, habituation, and response to a spatial change in rats following septal or medial frontal cortical damage. Behavioral neuroscience, 103(5), 1009-16.
[PubMed:2803548]
[WorldCat]
[DOI]
↑Norman, G., & Eacott, M.J. (2005).
Dissociable effects of lesions to the perirhinal cortex and the postrhinal cortex on memory for context and objects in rats. Behavioral neuroscience, 119(2), 557-66.
[PubMed:15839802]
[WorldCat]
[DOI]
↑ 5.05.15.2Ennaceur, A., & Delacour, J. (1988).
A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behavioural brain research, 31(1), 47-59.
[PubMed:3228475]
[WorldCat]
[DOI]
↑ 6.06.1Ian Q. Whishaw. Bryan Kolb. The Behavior of the Laboratory Rat: A Handbook with Tests. Oxford University Press, USA,1 edition:2004
↑ 7.07.17.27.3Dix, S.L., & Aggleton, J.P. (1999).
Extending the spontaneous preference test of recognition: evidence of object-location and object-context recognition. Behavioural brain research, 99(2), 191-200.
[PubMed:10512585]
[WorldCat]
[DOI]
↑ 8.08.1Thinus-Blanc, C., L. Bouzouba, K. Chaix, N. Chapuis, M. Durup, & B. Poucet A study of spatial parameters encoded during exploration in hamsters Journal of Experimental Psychology: Animal Behavior Processes:1987,13,418-427
↑Save, E., Poucet, B., Foreman, N., & Buhot, M.C. (1992).
Object exploration and reactions to spatial and nonspatial changes in hooded rats following damage to parietal cortex or hippocampal formation. Behavioral neuroscience, 106(3), 447-56.
[PubMed:1616611]
[WorldCat]
↑Uekita, T., & Okanoya, K. (2011).
Hippocampus lesions induced deficits in social and spatial recognition in Octodon degus. Behavioural brain research, 219(2), 302-9.
[PubMed:21291914]
[WorldCat]
[DOI]
↑ 12.012.1Mumby, D.G., Gaskin, S., Glenn, M.J., Schramek, T.E., & Lehmann, H. (2002).
Hippocampal damage and exploratory preferences in rats: memory for objects, places, and contexts. Learning & memory (Cold Spring Harbor, N.Y.), 9(2), 49-57.
[PubMed:11992015]
[PMC]
[WorldCat]
[DOI]
↑Norman, G., & Eacott, M.J. (2005).
Dissociable effects of lesions to the perirhinal cortex and the postrhinal cortex on memory for context and objects in rats. Behavioral neuroscience, 119(2), 557-66.
[PubMed:15839802]
[WorldCat]
[DOI]
↑Mishkin, M. (1978).
Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature, 273(5660), 297-8.
[PubMed:418358]
[WorldCat]
[DOI]
↑Murray, E.A., & Mishkin, M. (1998).
Object recognition and location memory in monkeys with excitotoxic lesions of the amygdala and hippocampus. The Journal of neuroscience : the official journal of the Society for Neuroscience, 18(16), 6568-82.
[PubMed:9698344]
[PMC]
[WorldCat]