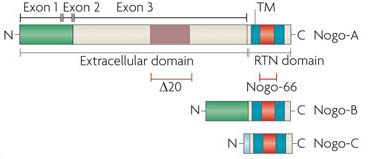
Reticulon-4, also known as Neurite outgrowth inhibitor or Nogo, is a protein that in humans is encoded by the RTN4 gene[1][2][3] that has been identified as an inhibitor of neurite outgrowth specific to the central nervous system.<br>This gene belongs to the family of reticulon-encoding genes. Reticulons are associated with the endoplasmic reticulum, and are involved in neuroendocrine secretion or in membrane trafficking in neuroendocrine cells. The product of this gene is a potent neurite outgrowth inhibitor that may also help block the regeneration of the central nervous system in higher vertebrates. Alternatively spliced transcript variants derived both from differential splicing and differential promoter usage and encoding different isoforms have been identified.[3] There are three isoforms: Nogo A, B and C. Nogo-A has two known inhibitory domains including amino-Nogo, at the N-terminus and Nogo-66, which makes up the molecules extracellular loop. Both amino-Nogo and Nogo-66 are involved in inhibitory responses, where amino-Nogo is a strong inhibitor of neurite outgrowth, and Nogo-66 is involved in growth cone destruction.[4]<br>Research suggests that blocking Nogo-A during neuronal damage (from diseases such as Multiple Sclerosis) will help to protect or restore the damaged neurons.[4] The investigation into the mechanisms of this protein presents a great potential for the treatment of auto-immune mediated demyelinating diseases and spinal cord injury regeneration. It has also been found to be a key player in the process whereby physical exercise enhances learning and memory processes in the brain.[5]
<br>
<br>
The membrane protein Nogo-A is a major player in the neurite growth-inhibitory and regeneration-inhibitory effects exerted by myelin in the mammalian brain and spinal cord. In the injured CNS, neutralization or blockade of Nogo-A enhances regeneration, compensatory fibre sprouting and functional recovery. In the intact nervous system, a number of physiological functions of Nogo proteins have been recently discovered, as discussed in this Review.<br>Nogo proteins and the related reticulon (RTN)1-3 proteins consist of a highly conserved, 200-amino acid carboxy-terminal RTN domain and non-homologous amino-terminal extensions of various lengths. The neurite growth-inhibitory protein Nogo-A appeared in evolution for the first time in frogs and is present in all higher vertebrates. Two active sites are present; a Nogo-A-specific domain and a 66-amino acid domain that lies between the transmembrane and intramembrane parts of the RTN domain. Nogo-A is highly enriched in the nervous system, in oligodendrocytes and myelin at adult stages, and in neurons and precursor cells during development. The short proteins Nogo-B and Nogo-C are not inhibitory and occur in various tissues, including the nervous system.<br>Two binding sites are currently known for the Nogo-66 sequence, the Nogo receptor 1 (NgR1) and the membrane protein paired immunoglobulin-like receptor B (PIRB). Both receptors also interact with other ligands, however. The receptor for the Nogo-A specific active site remains to be characterized. Rho activation followed by destabilizing effects on the cytoskeleton are obligatory steps in the postreceptor signalling and effector pathway that leads to the collapse of neurite growth cones. Several additional proteins are associated with what is probably a multisubunit receptor complex for Nogo-A.<br>Nogo-B, by interaction with a Nogo-B receptor (NGBR), influences vascular endothelial cells and smooth muscle cells, which hyperproliferate after vascular lesions in Nogo-A and Nogo-B double knockout mice. The function of Nogo-C is currently still unknown.<br>During CNS development, Nogo-A and its receptors are expressed in cortical precursors and affect their migration. Many projection neurons in the central and peripheral nervous systems express Nogo-A during axonal outgrowth; its neutralization or knockout enhances axonal fasciculation and influences branching. NgR1 and the shorter Nogo forms also have guidance and fasciculation functions in zebrafish, a lower vertebrate.<br>In the adult CNS, oligodendrocyte and myelin Nogo-A suppresses the growth programme of adult neurons, probably by a retrograde action on the cell bodies. Locally, neurite growth is dampened by the growth cone collapsing actions of Nogo-A. Nogo-A thus acts as a stabilizer of the adult CNS neuronal network and wiring. Ablation of Nogo-A or NgR1 accordingly enhances plastic rearrangements of CNS connections, extending the so-called 'critical period' far into adult ages, for example, for visual cortex plasticity. The schizophrenia-like behaviour of Nogo-A knockout mice and the associations found between psychiatric disorders and mutations in the genes encoding Nogo or NgR1 may be based on similar functions.<br>In addition to its cell surface expression, high amounts of Nogo are also present intracellularly. In neurons, its interaction with β-secretase points to a role in the regulation of amyloid precursor protein (APP) processing. Manipulations of Nogo have indicated a structural role for Nogo in the endoplasmic reticulum (ER) and the nuclear membrane. Interactions with proteins involved in cell survival and apoptosis have also been observed.<br>Various approaches aimed at suppressing Nogo-A or NgR1 actions have been used following injury of the adult spinal cord or brain. Acute functional suppression and, with more variable effects, chronic genetic deletion enhance regenerative sprouting and growth of various CNS tract systems. In addition, spared fibre systems have shown enhanced compensatory sprouting; both these processes were associated with substantial improvements of the behavioural recovery of lost functions in rodents and monkeys. These results illustrate the important growth-suppressive role of Nogo-A in the adult mammalian CNS.
Reticulon-4, also known as Neurite outgrowth inhibitor or Nogo, is a protein that in humans is encoded by the RTN4 gene[1][2][3] that has been identified as an inhibitor of neurite outgrowth specific to the central nervous system. This gene belongs to the family of reticulon-encoding genes. Reticulons are associated with the endoplasmic reticulum, and are involved in neuroendocrine secretion or in membrane trafficking in neuroendocrine cells. The product of this gene is a potent neurite outgrowth inhibitor that may also help block the regeneration of the central nervous system in higher vertebrates. Alternatively spliced transcript variants derived both from differential splicing and differential promoter usage and encoding different isoforms have been identified.[3] There are three isoforms: Nogo A, B and C. Nogo-A has two known inhibitory domains including amino-Nogo, at the N-terminus and Nogo-66, which makes up the molecules extracellular loop. Both amino-Nogo and Nogo-66 are involved in inhibitory responses, where amino-Nogo is a strong inhibitor of neurite outgrowth, and Nogo-66 is involved in growth cone destruction.[4] Research suggests that blocking Nogo-A during neuronal damage (from diseases such as Multiple Sclerosis) will help to protect or restore the damaged neurons.[4] The investigation into the mechanisms of this protein presents a great potential for the treatment of auto-immune mediated demyelinating diseases and spinal cord injury regeneration. It has also been found to be a key player in the process whereby physical exercise enhances learning and memory processes in the brain.[5]
The membrane protein Nogo-A is a major player in the neurite growth-inhibitory and regeneration-inhibitory effects exerted by myelin in the mammalian brain and spinal cord. In the injured CNS, neutralization or blockade of Nogo-A enhances regeneration, compensatory fibre sprouting and functional recovery. In the intact nervous system, a number of physiological functions of Nogo proteins have been recently discovered, as discussed in this Review. Nogo proteins and the related reticulon (RTN)1-3 proteins consist of a highly conserved, 200-amino acid carboxy-terminal RTN domain and non-homologous amino-terminal extensions of various lengths. The neurite growth-inhibitory protein Nogo-A appeared in evolution for the first time in frogs and is present in all higher vertebrates. Two active sites are present; a Nogo-A-specific domain and a 66-amino acid domain that lies between the transmembrane and intramembrane parts of the RTN domain. Nogo-A is highly enriched in the nervous system, in oligodendrocytes and myelin at adult stages, and in neurons and precursor cells during development. The short proteins Nogo-B and Nogo-C are not inhibitory and occur in various tissues, including the nervous system. Two binding sites are currently known for the Nogo-66 sequence, the Nogo receptor 1 (NgR1) and the membrane protein paired immunoglobulin-like receptor B (PIRB). Both receptors also interact with other ligands, however. The receptor for the Nogo-A specific active site remains to be characterized. Rho activation followed by destabilizing effects on the cytoskeleton are obligatory steps in the postreceptor signalling and effector pathway that leads to the collapse of neurite growth cones. Several additional proteins are associated with what is probably a multisubunit receptor complex for Nogo-A. Nogo-B, by interaction with a Nogo-B receptor (NGBR), influences vascular endothelial cells and smooth muscle cells, which hyperproliferate after vascular lesions in Nogo-A and Nogo-B double knockout mice. The function of Nogo-C is currently still unknown. During CNS development, Nogo-A and its receptors are expressed in cortical precursors and affect their migration. Many projection neurons in the central and peripheral nervous systems express Nogo-A during axonal outgrowth; its neutralization or knockout enhances axonal fasciculation and influences branching. NgR1 and the shorter Nogo forms also have guidance and fasciculation functions in zebrafish, a lower vertebrate. In the adult CNS, oligodendrocyte and myelin Nogo-A suppresses the growth programme of adult neurons, probably by a retrograde action on the cell bodies. Locally, neurite growth is dampened by the growth cone collapsing actions of Nogo-A. Nogo-A thus acts as a stabilizer of the adult CNS neuronal network and wiring. Ablation of Nogo-A or NgR1 accordingly enhances plastic rearrangements of CNS connections, extending the so-called 'critical period' far into adult ages, for example, for visual cortex plasticity. The schizophrenia-like behaviour of Nogo-A knockout mice and the associations found between psychiatric disorders and mutations in the genes encoding Nogo or NgR1 may be based on similar functions. In addition to its cell surface expression, high amounts of Nogo are also present intracellularly. In neurons, its interaction with β-secretase points to a role in the regulation of amyloid precursor protein (APP) processing. Manipulations of Nogo have indicated a structural role for Nogo in the endoplasmic reticulum (ER) and the nuclear membrane. Interactions with proteins involved in cell survival and apoptosis have also been observed. Various approaches aimed at suppressing Nogo-A or NgR1 actions have been used following injury of the adult spinal cord or brain. Acute functional suppression and, with more variable effects, chronic genetic deletion enhance regenerative sprouting and growth of various CNS tract systems. In addition, spared fibre systems have shown enhanced compensatory sprouting; both these processes were associated with substantial improvements of the behavioural recovery of lost functions in rodents and monkeys. These results illustrate the important growth-suppressive role of Nogo-A in the adult mammalian CNS.