今からおよそ80年前に、スペインの神経学者Ramon y Cajalが再生阻害の謎を解く重要なヒントを見いだした<ref name="1">Ramon y Cajal, S. Degeneration and regeneration of the nervous system. Hafner, New York, 1928.</ref>。彼は、感覚を伝える後根神経という末梢神経の軸索を切断し、その後の軸索の再生を観察した。再生しかけた軸索は、脊髄の中に侵入できず、再生できなかった。その後1980年代になって、Aguayoらは、脊髄の損傷による欠損部を末梢神経の周囲組織をグラフトとして移植することで、このグラフト内を軸索が再生する結果を得た。これらにより、神経細胞自体に再生する力がないのではなく、神経細胞を取り巻く環境が再生に適していないのではないかと考えられるに至った。<br> 1980年代、更に研究が進展し、ミエリンが神経突起の伸展を抑制することが報告された。そして、Schwabらは、ミエリンの中に再生を阻害している分子が存在していると考え、ミエリンの各フラクションに対する抗体を作成した。In vitroの実験により、IN-1抗体はミエリンの作用を中和し、220 kDaの糖蛋白に結合することが判明した。また、IN-1抗体を脊髄損傷させたラットに投与すると、軸索再生と運動機能の回復が認められた。これら一連の成果により、軸索再生阻害という概念は、仮説ではなく実在のものとして信じられるようになった。しかし再生阻害を担う蛋白の単離は10年かかっても実現しなかった。
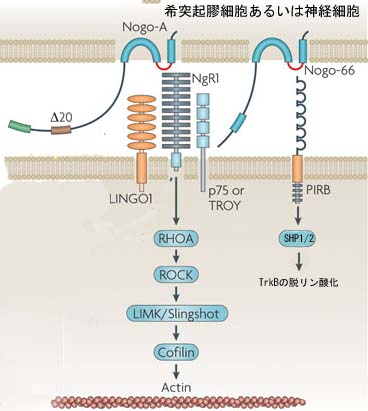
Two binding sites are currently known for the Nogo-66 sequence, the Nogo receptor 1 (NgR1) and the membrane protein paired immunoglobulin-like receptor B (PIRB). Both receptors also interact with other ligands, however. The receptor for the Nogo-A specific active site remains to be characterized. Rho activation followed by destabilizing effects on the cytoskeleton are obligatory steps in the postreceptor signalling and effector pathway that leads to the collapse of neurite growth cones. Several additional proteins are associated with what is probably a multisubunit receptor complex for Nogo-A.<br>Nogo-B, by interaction with a Nogo-B receptor (NGBR), influences vascular endothelial cells and smooth muscle cells, which hyperproliferate after vascular lesions in Nogo-A and Nogo-B double knockout mice. The function of Nogo-C is currently still unknown.<br>During CNS development, Nogo-A and its receptors are expressed in cortical precursors and affect their migration. Many projection neurons in the central and peripheral nervous systems express Nogo-A during axonal outgrowth; its neutralization or knockout enhances axonal fasciculation and influences branching. NgR1 and the shorter Nogo forms also have guidance and fasciculation functions in zebrafish, a lower vertebrate.<br>In the adult CNS, oligodendrocyte and myelin Nogo-A suppresses the growth programme of adult neurons, probably by a retrograde action on the cell bodies. Locally, neurite growth is dampened by the growth cone collapsing actions of Nogo-A. Nogo-A thus acts as a stabilizer of the adult CNS neuronal network and wiring. Ablation of Nogo-A or NgR1 accordingly enhances plastic rearrangements of CNS connections, extending the so-called 'critical period' far into adult ages, for example, for visual cortex plasticity. The schizophrenia-like behaviour of Nogo-A knockout mice and the associations found between psychiatric disorders and mutations in the genes encoding Nogo or NgR1 may be based on similar functions.<br>In addition to its cell surface expression, high amounts of Nogo are also present intracellularly. In neurons, its interaction with β-secretase points to a role in the regulation of amyloid precursor protein (APP) processing. Manipulations of Nogo have indicated a structural role for Nogo in the endoplasmic reticulum (ER) and the nuclear membrane. Interactions with proteins involved in cell survival and apoptosis have also been observed.<br>Various approaches aimed at suppressing Nogo-A or NgR1 actions have been used following injury of the adult spinal cord or brain. Acute functional suppression and, with more variable effects, chronic genetic deletion enhance regenerative sprouting and growth of various CNS tract systems. In addition, spared fibre systems have shown enhanced compensatory sprouting; both these processes were associated with substantial improvements of the behavioural recovery of lost functions in rodents and monkeys. These results illustrate the important growth-suppressive role of Nogo-A in the adult mammalian CNS.
Two binding sites are currently known for the Nogo-66 sequence, the Nogo receptor 1 (NgR1) and the membrane protein paired immunoglobulin-like receptor B (PIRB). Both receptors also interact with other ligands, however. The receptor for the Nogo-A specific active site remains to be characterized. Rho activation followed by destabilizing effects on the cytoskeleton are obligatory steps in the postreceptor signalling and effector pathway that leads to the collapse of neurite growth cones. Several additional proteins are associated with what is probably a multisubunit receptor complex for Nogo-A. Nogo-B, by interaction with a Nogo-B receptor (NGBR), influences vascular endothelial cells and smooth muscle cells, which hyperproliferate after vascular lesions in Nogo-A and Nogo-B double knockout mice. The function of Nogo-C is currently still unknown. During CNS development, Nogo-A and its receptors are expressed in cortical precursors and affect their migration. Many projection neurons in the central and peripheral nervous systems express Nogo-A during axonal outgrowth; its neutralization or knockout enhances axonal fasciculation and influences branching. NgR1 and the shorter Nogo forms also have guidance and fasciculation functions in zebrafish, a lower vertebrate. In the adult CNS, oligodendrocyte and myelin Nogo-A suppresses the growth programme of adult neurons, probably by a retrograde action on the cell bodies. Locally, neurite growth is dampened by the growth cone collapsing actions of Nogo-A. Nogo-A thus acts as a stabilizer of the adult CNS neuronal network and wiring. Ablation of Nogo-A or NgR1 accordingly enhances plastic rearrangements of CNS connections, extending the so-called 'critical period' far into adult ages, for example, for visual cortex plasticity. The schizophrenia-like behaviour of Nogo-A knockout mice and the associations found between psychiatric disorders and mutations in the genes encoding Nogo or NgR1 may be based on similar functions. In addition to its cell surface expression, high amounts of Nogo are also present intracellularly. In neurons, its interaction with β-secretase points to a role in the regulation of amyloid precursor protein (APP) processing. Manipulations of Nogo have indicated a structural role for Nogo in the endoplasmic reticulum (ER) and the nuclear membrane. Interactions with proteins involved in cell survival and apoptosis have also been observed. Various approaches aimed at suppressing Nogo-A or NgR1 actions have been used following injury of the adult spinal cord or brain. Acute functional suppression and, with more variable effects, chronic genetic deletion enhance regenerative sprouting and growth of various CNS tract systems. In addition, spared fibre systems have shown enhanced compensatory sprouting; both these processes were associated with substantial improvements of the behavioural recovery of lost functions in rodents and monkeys. These results illustrate the important growth-suppressive role of Nogo-A in the adult mammalian CNS.