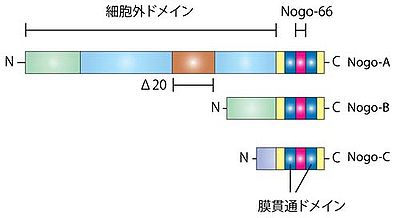
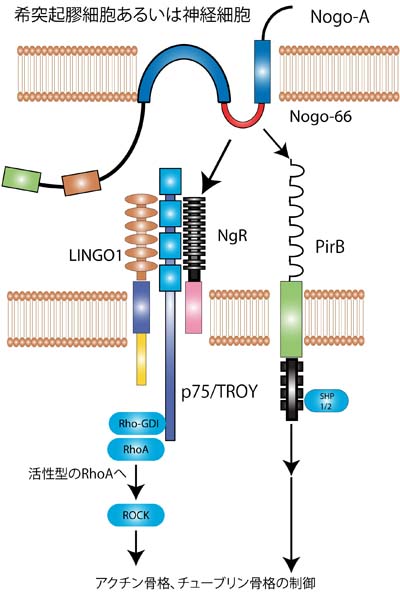
↑ 1.01.11.2Schwab, M.E. (2010).
Functions of Nogo proteins and their receptors in the nervous system. Nature reviews. Neuroscience, 11(12), 799-811.
[PubMed:21045861]
[WorldCat]
[DOI]
↑Schnell, L., & Schwab, M.E. (1990).
Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature, 343(6255), 269-72.
[PubMed:2300171]
[WorldCat]
[DOI]
↑Chen, M.S., Huber, A.B., van der Haar, M.E., Frank, M., Schnell, L., Spillmann, A.A., ..., & Schwab, M.E. (2000).
Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature, 403(6768), 434-9.
[PubMed:10667796]
[WorldCat]
[DOI]
↑GrandPré, T., Nakamura, F., Vartanian, T., & Strittmatter, S.M. (2000).
Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature, 403(6768), 439-44.
[PubMed:10667797]
[WorldCat]
[DOI]
↑Prinjha, R., Moore, S.E., Vinson, M., Blake, S., Morrow, R., Christie, G., ..., & Walsh, F.S. (2000).
Inhibitor of neurite outgrowth in humans. Nature, 403(6768), 383-4.
[PubMed:10667780]
[WorldCat]
[DOI]
↑Fournier, A.E., GrandPre, T., & Strittmatter, S.M. (2001).
Identification of a receptor mediating Nogo-66 inhibition of axonal regeneration. Nature, 409(6818), 341-6.
[PubMed:11201742]
[WorldCat]
[DOI]
↑Yamashita, T., Higuchi, H., & Tohyama, M. (2002).
The p75 receptor transduces the signal from myelin-associated glycoprotein to Rho. The Journal of cell biology, 157(4), 565-70.
[PubMed:12011108]
[PMC]
[WorldCat]
[DOI]
↑Wang, K.C., Kim, J.A., Sivasankaran, R., Segal, R., & He, Z. (2002).
P75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature, 420(6911), 74-8.
[PubMed:12422217]
[WorldCat]
[DOI]
↑Yamashita, T., & Tohyama, M. (2003).
The p75 receptor acts as a displacement factor that releases Rho from Rho-GDI. Nature neuroscience, 6(5), 461-7.
[PubMed:12692556]
[WorldCat]
[DOI]
↑Mi, S., Lee, X., Shao, Z., Thill, G., Ji, B., Relton, J., ..., & Pepinsky, R.B. (2004).
LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nature neuroscience, 7(3), 221-8.
[PubMed:14966521]
[WorldCat]
[DOI]
↑Atwal, J.K., Pinkston-Gosse, J., Syken, J., Stawicki, S., Wu, Y., Shatz, C., & Tessier-Lavigne, M. (2008).
PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science (New York, N.Y.), 322(5903), 967-70.
[PubMed:18988857]
[WorldCat]
[DOI]
↑Sato, Y., Iketani, M., Kurihara, Y., Yamaguchi, M., Yamashita, N., Nakamura, F., ..., & Takei, K. (2011).
Cartilage acidic protein-1B (LOTUS), an endogenous Nogo receptor antagonist for axon tract formation. Science (New York, N.Y.), 333(6043), 769-73.
[PubMed:21817055]
[PMC]
[WorldCat]
[DOI]