今からおよそ80年前のスペインの神経学者Ramon y Cajal、その後のAguayoらの実験により、神経細胞自体には再生する力があり、神経細胞を取り巻く環境が再生に適していないのではないかと考えられるようになる。その候補分子の一つとして、ミエリンが神経突起の伸展を抑制することが報告されたことから、ミエリンの中に再生を阻害し
↑Wang, K.C., Kim, J.A., Sivasankaran, R., Segal, R., & He, Z. (2002).
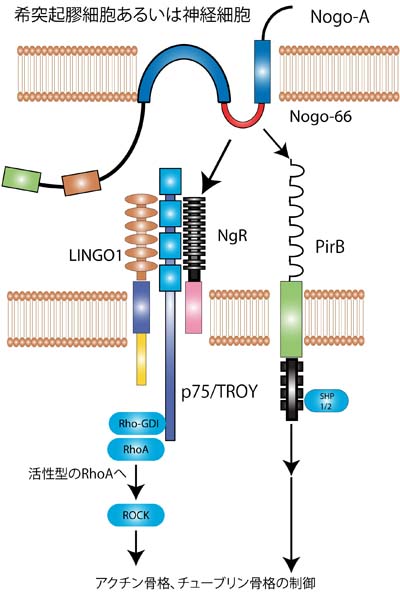
P75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature, 420(6911), 74-8.
[PubMed:12422217]
[WorldCat]
[DOI]
↑Mi, S., Lee, X., Shao, Z., Thill, G., Ji, B., Relton, J., ..., & Pepinsky, R.B. (2004).
LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nature neuroscience, 7(3), 221-8.
[PubMed:14966521]
[WorldCat]
[DOI]
↑Atwal, J.K., Pinkston-Gosse, J., Syken, J., Stawicki, S., Wu, Y., Shatz, C., & Tessier-Lavigne, M. (2008).
PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science (New York, N.Y.), 322(5903), 967-70.
[PubMed:18988857]
[WorldCat]
[DOI]
↑Schwab, M.E. (2010).
Functions of Nogo proteins and their receptors in the nervous system. Nature reviews. Neuroscience, 11(12), 799-811.
[PubMed:21045861]
[WorldCat]
[DOI]