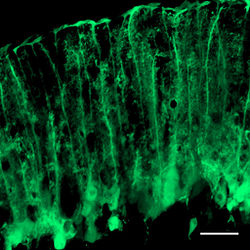
[[wikipedia:ja:サンティアゴ・ラモン・イ・カハール|Ramón y Cajal]]が1911年に出版した著名な解剖書中で、小脳皮質に柵状構造を形成する放射状線維を1850年代に最初に記載したBergmannに因み、「バーグマン線維をもつ上皮細胞」と表現したことから、バーグマングリアの名が使われるようになった。1885年に放射状線維がグリア細胞に由来することを見出した[[wikipedia:ja:カミッロ・ゴルジ|Camillo Golgi]]の名を取り、ゴルジ上皮細胞と呼ばれることもある。
[[wikipedia:Ramón y Cajal|Ramón y Cajal]]が1911年に出版した著名な解剖書中で、小脳皮質に柵状構造を形成する放射状線維を1850年代に最初に記載したBergmannに因み、「バーグマン線維をもつ上皮細胞」と表現したことから、バーグマングリアの名が使われるようになった。1885年に放射状線維が[[グリア細胞]]に由来することを見出した[[wikipedia:Camillo Golgi|Camillo Golgi]]の名を取り、ゴルジ上皮細胞と呼ばれることもある。
Ramón y Cajalが1911年に出版した著名な解剖書中で、小脳皮質に柵状構造を形成する放射状線維を1850年代に最初に記載したBergmannに因み、「バーグマン線維をもつ上皮細胞」と表現したことから、バーグマングリアの名が使われるようになった。1885年に放射状線維がグリア細胞に由来することを見出したCamillo Golgiの名を取り、ゴルジ上皮細胞と呼ばれることもある。
↑Sudarov, A., Turnbull, R.K., Kim, E.J., Lebel-Potter, M., Guillemot, F., & Joyner, A.L. (2011).
Ascl1 genetics reveals insights into cerebellum local circuit assembly. The Journal of neuroscience : the official journal of the Society for Neuroscience, 31(30), 11055-69.
[PubMed:21795554]
[PMC]
[WorldCat]
[DOI]
↑NARAHASHI, T., DEGUCHI, T., URAKAWA, N., & OHKUBO, Y. (1960).
Stabilization and rectification of muscle fiber membrane by tetrodotoxin. The American journal of physiology, 198, 934-8.
[PubMed:14426011]
[WorldCat]
[DOI]
↑NARAHASHI, T., DEGUCHI, T., URAKAWA, N., & OHKUBO, Y. (1960).
Stabilization and rectification of muscle fiber membrane by tetrodotoxin. The American journal of physiology, 198, 934-8.
[PubMed:14426011]
[WorldCat]
[DOI]
↑NARAHASHI, T., DEGUCHI, T., URAKAWA, N., & OHKUBO, Y. (1960).
Stabilization and rectification of muscle fiber membrane by tetrodotoxin. The American journal of physiology, 198, 934-8.
[PubMed:14426011]
[WorldCat]
[DOI]
↑NARAHASHI, T., DEGUCHI, T., URAKAWA, N., & OHKUBO, Y. (1960).
Stabilization and rectification of muscle fiber membrane by tetrodotoxin. The American journal of physiology, 198, 934-8.
[PubMed:14426011]
[WorldCat]
[DOI]
↑Eiraku, M., Tohgo, A., Ono, K., Kaneko, M., Fujishima, K., Hirano, T., & Kengaku, M. (2005).
DNER acts as a neuron-specific Notch ligand during Bergmann glial development. Nature neuroscience, 8(7), 873-80.
[PubMed:15965470]
[WorldCat]
[DOI]
↑Graus-Porta, D., Blaess, S., Senften, M., Littlewood-Evans, A., Damsky, C., Huang, Z., ..., & Müller, U. (2001).
Beta1-class integrins regulate the development of laminae and folia in the cerebral and cerebellar cortex. Neuron, 31(3), 367-79.
[PubMed:11516395]
[WorldCat]
[DOI]
↑Tanaka, M., Maeda, N., Noda, M., & Marunouchi, T. (2003).
A chondroitin sulfate proteoglycan PTPzeta /RPTPbeta regulates the morphogenesis of Purkinje cell dendrites in the developing cerebellum. The Journal of neuroscience : the official journal of the Society for Neuroscience, 23(7), 2804-14.
[PubMed:12684467]
[PMC]
[WorldCat]
↑Marcaggi, P., Billups, D., & Attwell, D. (2003).
The role of glial glutamate transporters in maintaining the independent operation of juvenile mouse cerebellar parallel fibre synapses. The Journal of physiology, 552(Pt 1), 89-107.
[PubMed:12878755]
[PMC]
[WorldCat]
[DOI]
↑Watase, K., Hashimoto, K., Kano, M., Yamada, K., Watanabe, M., Inoue, Y., ..., & Tanaka, K. (1998).
Motor discoordination and increased susceptibility to cerebellar injury in GLAST mutant mice. The European journal of neuroscience, 10(3), 976-88.
[PubMed:9753165]
[WorldCat]
[DOI]
↑Takayasu, Y., Iino, M., Shimamoto, K., Tanaka, K., & Ozawa, S. (2006).
Glial glutamate transporters maintain one-to-one relationship at the climbing fiber-Purkinje cell synapse by preventing glutamate spillover. The Journal of neuroscience : the official journal of the Society for Neuroscience, 26(24), 6563-72.
[PubMed:16775144]
[PMC]
[WorldCat]
[DOI]
↑Matsui, K., Jahr, C.E., & Rubio, M.E. (2005).
High-concentration rapid transients of glutamate mediate neural-glial communication via ectopic release. The Journal of neuroscience : the official journal of the Society for Neuroscience, 25(33), 7538-47.
[PubMed:16107641]
[PMC]
[WorldCat]
[DOI]
↑Riquelme, R., Miralles, C.P., & De Blas, A.L. (2002).
Bergmann glia GABA(A) receptors concentrate on the glial processes that wrap inhibitory synapses. The Journal of neuroscience : the official journal of the Society for Neuroscience, 22(24), 10720-30.
[PubMed:12486165]
[PMC]
[WorldCat]
↑Iino, M., Goto, K., Kakegawa, W., Okado, H., Sudo, M., Ishiuchi, S., ..., & Ozawa, S. (2001).
Glia-synapse interaction through Ca2+-permeable AMPA receptors in Bergmann glia. Science (New York, N.Y.), 292(5518), 926-9.
[PubMed:11340205]
[WorldCat]
[DOI]
↑Hoogland, T.M., Kuhn, B., Göbel, W., Huang, W., Nakai, J., Helmchen, F., ..., & Wang, S.S. (2009).
Radially expanding transglial calcium waves in the intact cerebellum. Proceedings of the National Academy of Sciences of the United States of America, 106(9), 3496-501.
[PubMed:19211787]
[PMC]
[WorldCat]
[DOI]
↑Shiwaku, H., Yoshimura, N., Tamura, T., Sone, M., Ogishima, S., Watase, K., ..., & Okazawa, H. (2010).
Suppression of the novel ER protein Maxer by mutant ataxin-1 in Bergman glia contributes to non-cell-autonomous toxicity. The EMBO journal, 29(14), 2446-60.
[PubMed:20531390]
[PMC]
[WorldCat]
[DOI]