↑ 1.01.1Carbone, E., & Lux, H.D. (1984).
A low voltage-activated, fully inactivating Ca channel in vertebrate sensory neurones. Nature, 310(5977), 501-2.
[PubMed:6087159]
[WorldCat]
[DOI]
↑Fedulova, S.A., Kostyuk, P.G., & Veselovsky, N.S. (1985).
Two types of calcium channels in the somatic membrane of new-born rat dorsal root ganglion neurones. The Journal of physiology, 359, 431-46.
[PubMed:2582115]
[PMC]
[WorldCat]
[DOI]
↑ 3.03.13.2Catterall, W.A., Perez-Reyes, E., Snutch, T.P., & Striessnig, J. (2005).
International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacological reviews, 57(4), 411-25.
[PubMed:16382099]
[WorldCat]
[DOI]
↑Hess, P., Lansman, J.B., & Tsien, R.W. (1984).
Different modes of Ca channel gating behaviour favoured by dihydropyridine Ca agonists and antagonists. Nature, 311(5986), 538-44.
[PubMed:6207437]
[WorldCat]
[DOI]
↑ 6.06.1Nowycky, M.C., Fox, A.P., & Tsien, R.W. (1985).
Three types of neuronal calcium channel with different calcium agonist sensitivity. Nature, 316(6027), 440-3.
[PubMed:2410796]
[WorldCat]
[DOI]
↑McCleskey, E.W., Fox, A.P., Feldman, D.H., Cruz, L.J., Olivera, B.M., Tsien, R.W., & Yoshikami, D. (1987).
Omega-conotoxin: direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proceedings of the National Academy of Sciences of the United States of America, 84(12), 4327-31.
[PubMed:2438698]
[PMC]
[WorldCat]
[DOI]
↑Llinás, R.R., Sugimori, M., & Cherksey, B. (1989).
Voltage-dependent calcium conductances in mammalian neurons. The P channel. Annals of the New York Academy of Sciences, 560, 103-11.
[PubMed:2545128]
[WorldCat]
[DOI]
↑Mintz, I.M., Adams, M.E., & Bean, B.P. (1992).
P-type calcium channels in rat central and peripheral neurons. Neuron, 9(1), 85-95.
[PubMed:1321648]
[WorldCat]
[DOI]
↑Randall, A., & Tsien, R.W. (1995).
Pharmacological dissection of multiple types of Ca2+ channel currents in rat cerebellar granule neurons. The Journal of neuroscience : the official journal of the Society for Neuroscience, 15(4), 2995-3012.
[PubMed:7722641]
[WorldCat]
↑Bourinet, E., Soong, T.W., Sutton, K., Slaymaker, S., Mathews, E., Monteil, A., ..., & Snutch, T.P. (1999).
Splicing of alpha 1A subunit gene generates phenotypic variants of P- and Q-type calcium channels. Nature neuroscience, 2(5), 407-15.
[PubMed:10321243]
[WorldCat]
[DOI]
↑Newcomb, R., Szoke, B., Palma, A., Wang, G., Chen, X.h., Hopkins, W., ..., & Miljanich, G. (1998).
Selective peptide antagonist of the class E calcium channel from the venom of the tarantula Hysterocrates gigas. Biochemistry, 37(44), 15353-62.
[PubMed:9799496]
[WorldCat]
[DOI]
↑Tanabe, T., Takeshima, H., Mikami, A., Flockerzi, V., Takahashi, H., Kangawa, K., ..., & Numa, S. (1987).
Primary structure of the receptor for calcium channel blockers from skeletal muscle. Nature, 328(6128), 313-8.
[PubMed:3037387]
[WorldCat]
[DOI]
↑Davies, A., Hendrich, J., Van Minh, A.T., Wratten, J., Douglas, L., & Dolphin, A.C. (2007).
Functional biology of the alpha(2)delta subunits of voltage-gated calcium channels. Trends in pharmacological sciences, 28(5), 220-8.
[PubMed:17403543]
[WorldCat]
[DOI]
↑Mori, Y., Friedrich, T., Kim, M.S., Mikami, A., Nakai, J., Ruth, P., ..., & Furuichi, T. (1991).
Primary structure and functional expression from complementary DNA of a brain calcium channel. Nature, 350(6317), 398-402.
[PubMed:1849233]
[WorldCat]
[DOI]
↑ 16.016.1Buraei, Z., & Yang, J. (2010).
The ß subunit of voltage-gated Ca2+ channels. Physiological reviews, 90(4), 1461-506.
[PubMed:20959621]
[PMC]
[WorldCat]
[DOI]
↑ 17.017.1Chen, R.S., Deng, T.C., Garcia, T., Sellers, Z.M., & Best, P.M. (2007).
Calcium channel gamma subunits: a functionally diverse protein family. Cell biochemistry and biophysics, 47(2), 178-86.
[PubMed:17652770]
[WorldCat]
↑Numa, S., Tanabe, T., Takeshima, H., Mikami, A., Niidome, T., Nishimura, S., ..., & Beam, K.G. (1990).
Molecular insights into excitation-contraction coupling. Cold Spring Harbor symposia on quantitative biology, 55, 1-7.
[PubMed:1966760]
[WorldCat]
[DOI]
↑Fabiato, A. (1983).
Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. The American journal of physiology, 245(1), C1-14.
[PubMed:6346892]
[WorldCat]
[DOI]
↑Eliasson, L., Abdulkader, F., Braun, M., Galvanovskis, J., Hoppa, M.B., & Rorsman, P. (2008).
Novel aspects of the molecular mechanisms controlling insulin secretion. The Journal of physiology, 586(14), 3313-24.
[PubMed:18511483]
[PMC]
[WorldCat]
[DOI]
↑Kollmar, R., Montgomery, L.G., Fak, J., Henry, L.J., & Hudspeth, A.J. (1997).
Predominance of the alpha1D subunit in L-type voltage-gated Ca2+ channels of hair cells in the chicken's cochlea. Proceedings of the National Academy of Sciences of the United States of America, 94(26), 14883-8.
[PubMed:9405708]
[PMC]
[WorldCat]
[DOI]
↑Schmitz, Y., & Witkovsky, P. (1997).
Dependence of photoreceptor glutamate release on a dihydropyridine-sensitive calcium channel. Neuroscience, 78(4), 1209-16.
[PubMed:9174087]
[WorldCat]
[DOI]
↑Bito, H., Deisseroth, K., & Tsien, R.W. (1996).
CREB phosphorylation and dephosphorylation: a Ca(2+)- and stimulus duration-dependent switch for hippocampal gene expression. Cell, 87(7), 1203-14.
[PubMed:8980227]
[WorldCat]
[DOI]
↑Takahashi, T., & Momiyama, A. (1993).
Different types of calcium channels mediate central synaptic transmission. Nature, 366(6451), 156-8.
[PubMed:7901765]
[WorldCat]
[DOI]
↑Wheeler, D.B., Randall, A., & Tsien, R.W. (1994).
Roles of N-type and Q-type Ca2+ channels in supporting hippocampal synaptic transmission. Science (New York, N.Y.), 264(5155), 107-11.
[PubMed:7832825]
[WorldCat]
[DOI]
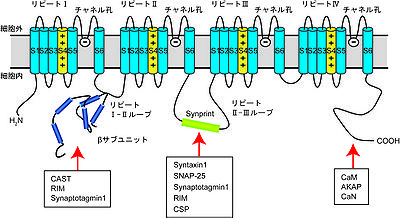
↑Kiyonaka, S., Nakajima, H., Takada, Y., Hida, Y., Yoshioka, T., Hagiwara, A., ..., & Ohtsuka, T. (2012).
Physical and functional interaction of the active zone protein CAST/ERC2 and the β-subunit of the voltage-dependent Ca(2+) channel. Journal of biochemistry, 152(2), 149-59.
[PubMed:22577167]
[WorldCat]
[DOI]
↑ 28.028.1Kiyonaka, S., Wakamori, M., Miki, T., Uriu, Y., Nonaka, M., Bito, H., ..., & Mori, Y. (2007).
RIM1 confers sustained activity and neurotransmitter vesicle anchoring to presynaptic Ca2+ channels. Nature neuroscience, 10(6), 691-701.
[PubMed:17496890]
[PMC]
[WorldCat]
[DOI]
↑Vendel, A.C., Terry, M.D., Striegel, A.R., Iverson, N.M., Leuranguer, V., Rithner, C.D., ..., & Horne, W.A. (2006).
Alternative splicing of the voltage-gated Ca2+ channel beta4 subunit creates a uniquely folded N-terminal protein binding domain with cell-specific expression in the cerebellar cortex. The Journal of neuroscience : the official journal of the Society for Neuroscience, 26(10), 2635-44.
[PubMed:16525042]
[PMC]
[WorldCat]
[DOI]
↑Wang, Y., Okamoto, M., Schmitz, F., Hofmann, K., & Südhof, T.C. (1997).
Rim is a putative Rab3 effector in regulating synaptic-vesicle fusion. Nature, 388(6642), 593-8.
[PubMed:9252191]
[WorldCat]
[DOI]
↑ 31.031.1Mangoni, M.E., Traboulsie, A., Leoni, A.L., Couette, B., Marger, L., Le Quang, K., ..., & Lory, P. (2006).
Bradycardia and slowing of the atrioventricular conduction in mice lacking CaV3.1/alpha1G T-type calcium channels. Circulation research, 98(11), 1422-30.
[PubMed:16690884]
[WorldCat]
[DOI]
↑Destexhe, A., Neubig, M., Ulrich, D., & Huguenard, J. (1998).
Dendritic low-threshold calcium currents in thalamic relay cells. The Journal of neuroscience : the official journal of the Society for Neuroscience, 18(10), 3574-88.
[PubMed:9570789]
[PMC]
[WorldCat]
↑Liu, L., Zwingman, T.A., & Fletcher, C.F. (2003).
In vivo analysis of voltage-dependent calcium channels. Journal of bioenergetics and biomembranes, 35(6), 671-85.
[PubMed:15000527]
[WorldCat]
[DOI]
↑Huc, S., Monteil, A., Bidaud, I., Barbara, G., Chemin, J., & Lory, P. (2009).
Regulation of T-type calcium channels: signalling pathways and functional implications. Biochimica et biophysica acta, 1793(6), 947-52.
[PubMed:19071165]
[WorldCat]
[DOI]
↑Montell, C., & Rubin, G.M. (1989).
Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron, 2(4), 1313-23.
[PubMed:2516726]
[WorldCat]
[DOI]
↑ 36.036.1Numata, T., Kozai, D., Takahashi, N., Kato, K., Uriu, Y., Yamamoto, S., ..., & Mori, Y. (2009).
[Structures and variable functions of TRP channels]. Seikagaku. The Journal of Japanese Biochemical Society, 81(11), 962-83.
[PubMed:19999578]
[WorldCat]