今からおよそ80年前のスペインの神経学者Ramon y Cajal、その後のAguayoらの実験により、神経細胞自体には再生する力があり、神経細胞を取り巻く環境が再生に適していないのではないかと考えられるようになる。その候補分子の一つとして、ミエリンが神経突起の伸展を抑制することが報告されたことから、ミエリンの中に再生を阻害している分子が存在していると考えられた。そして、Schwabらにより、ミエリンの各フラクションに対する抗体が作成され、IN-1抗体が発見される。[2]IN-1はミエリンの作用を打ち消し、また、IN-1抗体を脊髄損傷させたラットに投与すると、軸索再生と運動機能の回復が認められることが報告された。その後、3つのグループによりIN-1抗体の認識するペプチド配列をもとに、目的の蛋白がクローニングされ、Nogoと名付けられた。 [3][4][5]
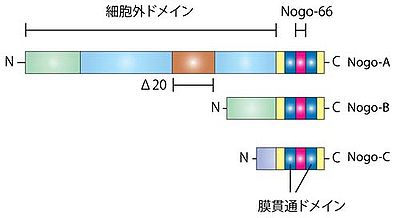
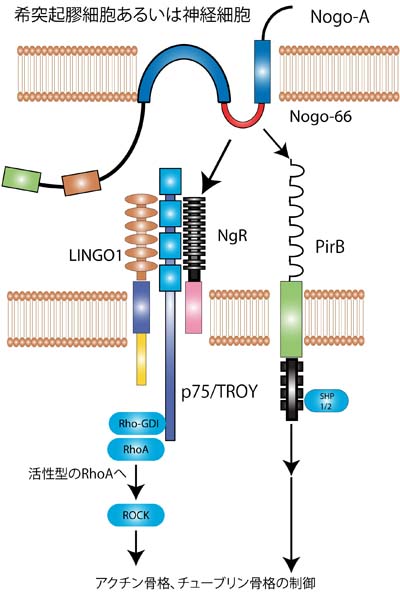
Nogo蛋白は培養神経細胞に対して突起伸展抑制作用及び軸索の先端の成長円錐を虚脱させる作用を持っており、特に、Δ20ドメインとNogo-66がその作用を担うことが報告された。
↑ 1.01.11.2Schwab, M.E. (2010).
Functions of Nogo proteins and their receptors in the nervous system. Nature reviews. Neuroscience, 11(12), 799-811.
[PubMed:21045861]
[WorldCat]
[DOI]
↑Schnell, L., & Schwab, M.E. (1990).
Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature, 343(6255), 269-72.
[PubMed:2300171]
[WorldCat]
[DOI]
↑Chen, M.S., Huber, A.B., van der Haar, M.E., Frank, M., Schnell, L., Spillmann, A.A., ..., & Schwab, M.E. (2000).
Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature, 403(6768), 434-9.
[PubMed:10667796]
[WorldCat]
[DOI]
↑GrandPré, T., Nakamura, F., Vartanian, T., & Strittmatter, S.M. (2000).
Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature, 403(6768), 439-44.
[PubMed:10667797]
[WorldCat]
[DOI]
↑Prinjha, R., Moore, S.E., Vinson, M., Blake, S., Morrow, R., Christie, G., ..., & Walsh, F.S. (2000).
Inhibitor of neurite outgrowth in humans. Nature, 403(6768), 383-4.
[PubMed:10667780]
[WorldCat]
[DOI]
↑Fournier, A.E., GrandPre, T., & Strittmatter, S.M. (2001).
Identification of a receptor mediating Nogo-66 inhibition of axonal regeneration. Nature, 409(6818), 341-6.
[PubMed:11201742]
[WorldCat]
[DOI]
↑Wang, K.C., Kim, J.A., Sivasankaran, R., Segal, R., & He, Z. (2002).
P75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature, 420(6911), 74-8.
[PubMed:12422217]
[WorldCat]
[DOI]
↑Yamashita, T., Higuchi, H., & Tohyama, M. (2002).
The p75 receptor transduces the signal from myelin-associated glycoprotein to Rho. The Journal of cell biology, 157(4), 565-70.
[PubMed:12011108]
[PMC]
[WorldCat]
[DOI]
↑Yamashita, T., & Tohyama, M. (2003).
The p75 receptor acts as a displacement factor that releases Rho from Rho-GDI. Nature neuroscience, 6(5), 461-7.
[PubMed:12692556]
[WorldCat]
[DOI]
↑Mi, S., Lee, X., Shao, Z., Thill, G., Ji, B., Relton, J., ..., & Pepinsky, R.B. (2004).
LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nature neuroscience, 7(3), 221-8.
[PubMed:14966521]
[WorldCat]
[DOI]
↑Atwal, J.K., Pinkston-Gosse, J., Syken, J., Stawicki, S., Wu, Y., Shatz, C., & Tessier-Lavigne, M. (2008).
PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science (New York, N.Y.), 322(5903), 967-70.
[PubMed:18988857]
[WorldCat]
[DOI]