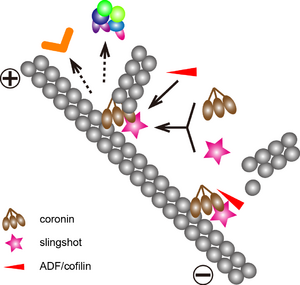
CoroninのArp2/3複合体への結合は、protein kinase C (PKC) によるリン酸化によって抑制される[48]。また、同部位は、SSHによる脱リン酸化制御も受ける[18]。CoroninはSSHのアクチンへの結合を媒介するため、coroninの制御によって、ADF/cofilinの活性が調節される。実際、coroninのノックダウンによって、細胞内のリン酸化cofilin量が増加する[18]。
↑ 1.01.11.21.3Bugyi, B., & Carlier, M.F. (2010).
Control of actin filament treadmilling in cell motility. Annual review of biophysics, 39, 449-70.
[PubMed:20192778]
[WorldCat]
[DOI]
↑ 2.02.12.22.3Medeiros, N.A., Burnette, D.T., & Forscher, P. (2006).
Myosin II functions in actin-bundle turnover in neuronal growth cones. Nature cell biology, 8(3), 215-26.
[PubMed:16501565]
[WorldCat]
[DOI]
↑ 3.03.1Mogilner, A., & Oster, G. (2003).
Polymer motors: pushing out the front and pulling up the back. Current biology : CB, 13(18), R721-33.
[PubMed:13678614]
[WorldCat]
[DOI]
↑ 4.04.14.2Van Goor, D., Hyland, C., Schaefer, A.W., & Forscher, P. (2012).
The role of actin turnover in retrograde actin network flow in neuronal growth cones. PloS one, 7(2), e30959.
[PubMed:22359556]
[PMC]
[WorldCat]
[DOI]
↑ 5.05.1Craig, E.M., Van Goor, D., Forscher, P., & Mogilner, A. (2012).
Membrane tension, myosin force, and actin turnover maintain actin treadmill in the nerve growth cone. Biophysical journal, 102(7), 1503-13.
[PubMed:22500750]
[PMC]
[WorldCat]
[DOI]
↑ 6.06.1Kiuchi, T., Nagai, T., Ohashi, K., & Mizuno, K. (2011).
Measurements of spatiotemporal changes in G-actin concentration reveal its effect on stimulus-induced actin assembly and lamellipodium extension. The Journal of cell biology, 193(2), 365-80.
[PubMed:21502360]
[PMC]
[WorldCat]
[DOI]
↑Mejillano, M.R., Kojima, S., Applewhite, D.A., Gertler, F.B., Svitkina, T.M., & Borisy, G.G. (2004).
Lamellipodial versus filopodial mode of the actin nanomachinery: pivotal role of the filament barbed end. Cell, 118(3), 363-73.
[PubMed:15294161]
[WorldCat]
[DOI]
↑Carlier, M.F., & Pantaloni, D. (1997).
Control of actin dynamics in cell motility. Journal of molecular biology, 269(4), 459-67.
[PubMed:9217250]
[WorldCat]
[DOI]
↑Akin, O., & Mullins, R.D. (2008).
Capping protein increases the rate of actin-based motility by promoting filament nucleation by the Arp2/3 complex. Cell, 133(5), 841-51.
[PubMed:18510928]
[PMC]
[WorldCat]
[DOI]
↑ 11.011.1Suraneni, P., Rubinstein, B., Unruh, J.R., Durnin, M., Hanein, D., & Li, R. (2012).
The Arp2/3 complex is required for lamellipodia extension and directional fibroblast cell migration. The Journal of cell biology, 197(2), 239-51.
[PubMed:22492726]
[PMC]
[WorldCat]
[DOI]
↑Strasser, G.A., Rahim, N.A., VanderWaal, K.E., Gertler, F.B., & Lanier, L.M. (2004).
Arp2/3 is a negative regulator of growth cone translocation. Neuron, 43(1), 81-94.
[PubMed:15233919]
[WorldCat]
[DOI]
↑Weaver, A.M., Heuser, J.E., Karginov, A.V., Lee, W.L., Parsons, J.T., & Cooper, J.A. (2002).
Interaction of cortactin and N-WASp with Arp2/3 complex. Current biology : CB, 12(15), 1270-8.
[PubMed:12176354]
[WorldCat]
[DOI]
↑Uruno, T., Liu, J., Li, Y., Smith, N., & Zhan, X. (2003).
Sequential interaction of actin-related proteins 2 and 3 (Arp2/3) complex with neural Wiscott-Aldrich syndrome protein (N-WASP) and cortactin during branched actin filament network formation. The Journal of biological chemistry, 278(28), 26086-93.
[PubMed:12732638]
[WorldCat]
[DOI]
↑dos Remedios, C.G., Chhabra, D., Kekic, M., Dedova, I.V., Tsubakihara, M., Berry, D.A., & Nosworthy, N.J. (2003).
Actin binding proteins: regulation of cytoskeletal microfilaments. Physiological reviews, 83(2), 433-73.
[PubMed:12663865]
[WorldCat]
[DOI]
↑Chan, C., Beltzner, C.C., & Pollard, T.D. (2009).
Cofilin dissociates Arp2/3 complex and branches from actin filaments. Current biology : CB, 19(7), 537-45.
[PubMed:19362000]
[PMC]
[WorldCat]
[DOI]
↑ 18.018.118.218.3Cai, L., Marshall, T.W., Uetrecht, A.C., Schafer, D.A., & Bear, J.E. (2007).
Coronin 1B coordinates Arp2/3 complex and cofilin activities at the leading edge. Cell, 128(5), 915-29.
[PubMed:17350576]
[PMC]
[WorldCat]
[DOI]
↑ 19.019.1Dayel, M.J., & Mullins, R.D. (2004).
Activation of Arp2/3 complex: addition of the first subunit of the new filament by a WASP protein triggers rapid ATP hydrolysis on Arp2. PLoS biology, 2(4), E91.
[PubMed:15094799]
[PMC]
[WorldCat]
[DOI]
↑Martin, A.C., Welch, M.D., & Drubin, D.G. (2006).
Arp2/3 ATP hydrolysis-catalysed branch dissociation is critical for endocytic force generation. Nature cell biology, 8(8), 826-33.
[PubMed:16862144]
[WorldCat]
[DOI]
↑Suter, D.M., & Forscher, P. (2001).
Transmission of growth cone traction force through apCAM-cytoskeletal linkages is regulated by Src family tyrosine kinase activity. The Journal of cell biology, 155(3), 427-38.
[PubMed:11673478]
[PMC]
[WorldCat]
[DOI]
↑Gardel, M.L., Sabass, B., Ji, L., Danuser, G., Schwarz, U.S., & Waterman, C.M. (2008).
Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of cell biology, 183(6), 999-1005.
[PubMed:19075110]
[PMC]
[WorldCat]
[DOI]
↑ 23.023.1Lai, F.P., Szczodrak, M., Block, J., Faix, J., Breitsprecher, D., Mannherz, H.G., ..., & Rottner, K. (2008).
Arp2/3 complex interactions and actin network turnover in lamellipodia. The EMBO journal, 27(7), 982-92.
[PubMed:18309290]
[PMC]
[WorldCat]
[DOI]
↑Nakagawa, H., Miki, H., Ito, M., Ohashi, K., Takenawa, T., & Miyamoto, S. (2001).
N-WASP, WAVE and Mena play different roles in the organization of actin cytoskeleton in lamellipodia. Journal of cell science, 114(Pt 8), 1555-65.
[PubMed:11282031]
[WorldCat]
↑Keren, K., Yam, P.T., Kinkhabwala, A., Mogilner, A., & Theriot, J.A. (2009).
Intracellular fluid flow in rapidly moving cells. Nature cell biology, 11(10), 1219-24.
[PubMed:19767741]
[PMC]
[WorldCat]
[DOI]
↑Dent, E.W., Gupton, S.L., & Gertler, F.B. (2011).
The growth cone cytoskeleton in axon outgrowth and guidance. Cold Spring Harbor perspectives in biology, 3(3).
[PubMed:21106647]
[PMC]
[WorldCat]
[DOI]
↑Yao, J., Sasaki, Y., Wen, Z., Bassell, G.J., & Zheng, J.Q. (2006).
An essential role for beta-actin mRNA localization and translation in Ca2+-dependent growth cone guidance. Nature neuroscience, 9(10), 1265-73.
[PubMed:16980965]
[WorldCat]
[DOI]
↑Leung, K.M., van Horck, F.P., Lin, A.C., Allison, R., Standart, N., & Holt, C.E. (2006).
Asymmetrical beta-actin mRNA translation in growth cones mediates attractive turning to netrin-1. Nature neuroscience, 9(10), 1247-56.
[PubMed:16980963]
[PMC]
[WorldCat]
[DOI]
↑Yang, N., Higuchi, O., Ohashi, K., Nagata, K., Wada, A., Kangawa, K., ..., & Mizuno, K. (1998).
Cofilin phosphorylation by LIM-kinase 1 and its role in Rac-mediated actin reorganization. Nature, 393(6687), 809-12.
[PubMed:9655398]
[WorldCat]
[DOI]
↑Niwa, R., Nagata-Ohashi, K., Takeichi, M., Mizuno, K., & Uemura, T. (2002).
Control of actin reorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell, 108(2), 233-46.
[PubMed:11832213]
[WorldCat]
[DOI]
↑Agnew, B.J., Minamide, L.S., & Bamburg, J.R. (1995).
Reactivation of phosphorylated actin depolymerizing factor and identification of the regulatory site. The Journal of biological chemistry, 270(29), 17582-7.
[PubMed:7615564]
[WorldCat]
[DOI]
↑ 32.032.1Bernstein, B.W., & Bamburg, J.R. (2010).
ADF/cofilin: a functional node in cell biology. Trends in cell biology, 20(4), 187-95.
[PubMed:20133134]
[PMC]
[WorldCat]
[DOI]
↑Maekawa, M., Ishizaki, T., Boku, S., Watanabe, N., Fujita, A., Iwamatsu, A., ..., & Narumiya, S. (1999).
Signaling from Rho to the actin cytoskeleton through protein kinases ROCK and LIM-kinase. Science (New York, N.Y.), 285(5429), 895-8.
[PubMed:10436159]
[WorldCat]
[DOI]
↑Ohashi, K., Nagata, K., Maekawa, M., Ishizaki, T., Narumiya, S., & Mizuno, K. (2000).
Rho-associated kinase ROCK activates LIM-kinase 1 by phosphorylation at threonine 508 within the activation loop. The Journal of biological chemistry, 275(5), 3577-82.
[PubMed:10652353]
[WorldCat]
[DOI]
↑Eiseler, T., Döppler, H., Yan, I.K., Kitatani, K., Mizuno, K., & Storz, P. (2009).
Protein kinase D1 regulates cofilin-mediated F-actin reorganization and cell motility through slingshot. Nature cell biology, 11(5), 545-56.
[PubMed:19329994]
[PMC]
[WorldCat]
[DOI]
↑Le Clainche, C., & Carlier, M.F. (2008).
Regulation of actin assembly associated with protrusion and adhesion in cell migration. Physiological reviews, 88(2), 489-513.
[PubMed:18391171]
[WorldCat]
[DOI]
↑ 38.038.1Vitriol, E.A., & Zheng, J.Q. (2012).
Growth cone travel in space and time: the cellular ensemble of cytoskeleton, adhesion, and membrane. Neuron, 73(6), 1068-81.
[PubMed:22445336]
[PMC]
[WorldCat]
[DOI]
↑Wen, Z., Han, L., Bamburg, J.R., Shim, S., Ming, G.L., & Zheng, J.Q. (2007).
BMP gradients steer nerve growth cones by a balancing act of LIM kinase and Slingshot phosphatase on ADF/cofilin. The Journal of cell biology, 178(1), 107-19.
[PubMed:17606869]
[PMC]
[WorldCat]
[DOI]
↑Takenawa, T., & Suetsugu, S. (2007).
The WASP-WAVE protein network: connecting the membrane to the cytoskeleton. Nature reviews. Molecular cell biology, 8(1), 37-48.
[PubMed:17183359]
[WorldCat]
[DOI]
↑Oikawa, T., Yamaguchi, H., Itoh, T., Kato, M., Ijuin, T., Yamazaki, D., ..., & Takenawa, T. (2004).
PtdIns(3,4,5)P3 binding is necessary for WAVE2-induced formation of lamellipodia. Nature cell biology, 6(5), 420-6.
[PubMed:15107862]
[WorldCat]
[DOI]
↑Shekarabi, M., Moore, S.W., Tritsch, N.X., Morris, S.J., Bouchard, J.F., & Kennedy, T.E. (2005).
Deleted in colorectal cancer binding netrin-1 mediates cell substrate adhesion and recruits Cdc42, Rac1, Pak1, and N-WASP into an intracellular signaling complex that promotes growth cone expansion. The Journal of neuroscience : the official journal of the Society for Neuroscience, 25(12), 3132-41.
[PubMed:15788770]
[PMC]
[WorldCat]
[DOI]
↑Myers, J.P., Robles, E., Ducharme-Smith, A., & Gomez, T.M. (2012).
Focal adhesion kinase modulates Cdc42 activity downstream of positive and negative axon guidance cues. Journal of cell science, 125(Pt 12), 2918-29.
[PubMed:22393238]
[PMC]
[WorldCat]
[DOI]
↑Kelley, L.C., Hayes, K.E., Ammer, A.G., Martin, K.H., & Weed, S.A. (2010).
Cortactin phosphorylated by ERK1/2 localizes to sites of dynamic actin regulation and is required for carcinoma lamellipodia persistence. PloS one, 5(11), e13847.
[PubMed:21079800]
[PMC]
[WorldCat]
[DOI]
↑Huang, C., Ni, Y., Wang, T., Gao, Y., Haudenschild, C.C., & Zhan, X. (1997).
Down-regulation of the filamentous actin cross-linking activity of cortactin by Src-mediated tyrosine phosphorylation. The Journal of biological chemistry, 272(21), 13911-5.
[PubMed:9153252]
[WorldCat]
[DOI]
↑Knöll, B., & Drescher, U. (2004).
Src family kinases are involved in EphA receptor-mediated retinal axon guidance. The Journal of neuroscience : the official journal of the Society for Neuroscience, 24(28), 6248-57.
[PubMed:15254079]
[PMC]
[WorldCat]
[DOI]
↑Cai, L., Holoweckyj, N., Schaller, M.D., & Bear, J.E. (2005).
Phosphorylation of coronin 1B by protein kinase C regulates interaction with Arp2/3 and cell motility. The Journal of biological chemistry, 280(36), 31913-23.
[PubMed:16027158]
[WorldCat]
[DOI]
↑Vicente-Manzanares, M., Ma, X., Adelstein, R.S., & Horwitz, A.R. (2009).
Non-muscle myosin II takes centre stage in cell adhesion and migration. Nature reviews. Molecular cell biology, 10(11), 778-90.
[PubMed:19851336]
[PMC]
[WorldCat]
[DOI]
↑Gallo, G. (2006).
RhoA-kinase coordinates F-actin organization and myosin II activity during semaphorin-3A-induced axon retraction. Journal of cell science, 119(Pt 16), 3413-23.
[PubMed:16899819]
[PMC]
[WorldCat]
[DOI]
↑Kubo, T., Endo, M., Hata, K., Taniguchi, J., Kitajo, K., Tomura, S., ..., & Yamashita, T. (2008).
Myosin IIA is required for neurite outgrowth inhibition produced by repulsive guidance molecule. Journal of neurochemistry, 105(1), 113-26.
[PubMed:18005226]
[WorldCat]
[DOI]
↑Murray, A., Naeem, A., Barnes, S.H., Drescher, U., & Guthrie, S. (2010).
Slit and Netrin-1 guide cranial motor axon pathfinding via Rho-kinase, myosin light chain kinase and myosin II. Neural development, 5, 16.
[PubMed:20569485]
[PMC]
[WorldCat]
[DOI]