脳においてノルアドレナリンの多くは、MAO、[[アルデヒド還元酵素]]、およびCOMTにより[[wikipedia:3-Methoxy-4-hydroxyphenylglycol|3-メトキシ-4-ヒドロキシフェニルグリコール]] (3-methoxy-4-hydroxyphenylglycol, MHPG)へ代謝され、さらに[[wikipedia:Vanillylmandelic acid|3-メトキシ-4-ヒドロキシマンデル酸]] (3-methoxy-4-hydroxymandelic acid) (または[[wikipedia:Vanillylmandelic acid|バニリルマンデル酸]] (vanillylmandelic acid, VMA)となって尿中に排出される<ref name="ref15">'''D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong'''<br> Principles of Pharmacology, Second Edition<br>''Wolters Kluwer Health (Philadelphia)'':2002</ref>。MHPGの硫酸化物も尿中に排出される<ref name="ref15" />。
== 受容体 ==
脳においてノルアドレナリンの多くは、MAO、[[アルデヒド還元酵素]]、およびCOMTにより[[wikipedia:3-Methoxy-4-hydroxyphenylglycol|3-メトキシ-4-ヒドロキシフェニルグリコール]] (3-methoxy-4-hydroxyphenylglycol, MHPG)へ代謝され、さらに[[wikipedia:Vanillylmandelic acid|3-メトキシ-4-ヒドロキシマンデル酸]] (3-methoxy-4-hydroxymandelic acid) (または[[wikipedia:Vanillylmandelic acid|バニリルマンデル酸]] (vanillylmandelic acid, VMA)となって尿中に排出される<ref name="ref15">'''D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong'''<br> Principles of Pharmacology, Second Edition<br>''Wolters Kluwer Health (Philadelphia)'':2002</ref>。MHPGの硫酸化物も尿中に排出される<ref name="ref15" />。
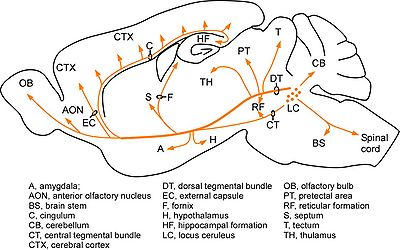
A6:青斑核(locus ceruleus)と呼ばれる。橋の背側に位置し、最も主要なノルアドレナリン作動性神経細胞の核である。青斑核からは、大脳皮質、視床、視床下部、小脳、中脳、脊髄、など脳のほぼ全域にわたって投射している(図2:Moore and Bloom, 1979[32]より許可を得て改変)。
↑U S von Euler A Specific Sympathomimetic Ergone in Adrenergic Nerve Fibres (Sympathin) and its Relations to Adrenaline and Nor-Adrenaline Acta Physiol., Scand. :1946, 12; 73–97
↑P Holtz Über die sympathicomimetische Wirksamkeit von Gehirnextrakten. Acta Physiol., Scand. : 1950, 20; 354–362
↑Dunkley, P.R., Bobrovskaya, L., Graham, M.E., von Nagy-Felsobuki, E.I., & Dickson, P.W. (2004).
Tyrosine hydroxylase phosphorylation: regulation and consequences. Journal of neurochemistry, 91(5), 1025-43.
[PubMed:15569247]
[WorldCat]
[DOI]
↑Daubner, S.C., Le, T., & Wang, S. (2011).
Tyrosine hydroxylase and regulation of dopamine synthesis. Archives of biochemistry and biophysics, 508(1), 1-12.
[PubMed:21176768]
[PMC]
[WorldCat]
[DOI]
↑Nagatsu, T. (1989).
The human tyrosine hydroxylase gene. Cellular and molecular neurobiology, 9(3), 313-21.
[PubMed:2575455]
[WorldCat]
[DOI]
↑Thöny, B., Auerbach, G., & Blau, N. (2000).
Tetrahydrobiopterin biosynthesis, regeneration and functions. The Biochemical journal, 347 Pt 1, 1-16.
[PubMed:10727395]
[PMC]
[WorldCat]
↑Rush, R.A., & Geffen, L.B. (1980).
Dopamine beta-hydroxylase in health and disease. Critical reviews in clinical laboratory sciences, 12(3), 241-77.
[PubMed:6998654]
[WorldCat]
[DOI]
↑Erickson, J.D., & Varoqui, H. (2000).
Molecular analysis of vesicular amine transporter function and targeting to secretory organelles. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 14(15), 2450-8.
[PubMed:11099462]
[WorldCat]
[DOI]
↑Mandela, P., & Ordway, G.A. (2006).
The norepinephrine transporter and its regulation. Journal of neurochemistry, 97(2), 310-33.
[PubMed:16539676]
[WorldCat]
[DOI]
↑ 14.014.1Youdim, M.B., Edmondson, D., & Tipton, K.F. (2006).
The therapeutic potential of monoamine oxidase inhibitors. Nature reviews. Neuroscience, 7(4), 295-309.
[PubMed:16552415]
[WorldCat]
[DOI]
↑Vitalis, T., Fouquet, C., Alvarez, C., Seif, I., Price, D., Gaspar, P., & Cases, O. (2002).
Developmental expression of monoamine oxidases A and B in the central and peripheral nervous systems of the mouse. The Journal of comparative neurology, 442(4), 331-47.
[PubMed:11793338]
[WorldCat]
[DOI]
↑Chen, J., Song, J., Yuan, P., Tian, Q., Ji, Y., Ren-Patterson, R., ..., & Weinberger, D.R. (2011).
Orientation and cellular distribution of membrane-bound catechol-O-methyltransferase in cortical neurons: implications for drug development. The Journal of biological chemistry, 286(40), 34752-60.
[PubMed:21846718]
[PMC]
[WorldCat]
[DOI]
↑ 17.017.117.217.3D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong Principles of Pharmacology, Second Edition Wolters Kluwer Health (Philadelphia):2002
↑Madison, D.V., & Nicoll, R.A. (1986).
Actions of noradrenaline recorded intracellularly in rat hippocampal CA1 pyramidal neurones, in vitro. The Journal of physiology, 372, 221-44.
[PubMed:2873241]
[PMC]
[WorldCat]
[DOI]
↑Haas, H.L., & Konnerth, A. (1983).
Histamine and noradrenaline decrease calcium-activated potassium conductance in hippocampal pyramidal cells. Nature, 302(5907), 432-4.
[PubMed:6300681]
[WorldCat]
[DOI]
↑Pedarzani, P., & Storm, J.F. (1993).
PKA mediates the effects of monoamine transmitters on the K+ current underlying the slow spike frequency adaptation in hippocampal neurons. Neuron, 11(6), 1023-35.
[PubMed:8274274]
[WorldCat]
[DOI]
↑Neuman, R.S., & Harley, C.W. (1983).
Long-lasting potentiation of the dentate gyrus population spike by norepinephrine. Brain research, 273(1), 162-5.
[PubMed:6311345]
[WorldCat]
[DOI]
↑O'Dell, T.J., Connor, S.A., Gelinas, J.N., & Nguyen, P.V. (2010).
Viagra for your synapses: Enhancement of hippocampal long-term potentiation by activation of beta-adrenergic receptors. Cellular signalling, 22(5), 728-36.
[PubMed:20043991]
[PMC]
[WorldCat]
[DOI]
↑Hoffman, D.A., & Johnston, D. (1999).
Neuromodulation of dendritic action potentials. Journal of neurophysiology, 81(1), 408-11.
[PubMed:9914302]
[WorldCat]
[DOI]
↑Yuan, L.L., Adams, J.P., Swank, M., Sweatt, J.D., & Johnston, D. (2002).
Protein kinase modulation of dendritic K+ channels in hippocampus involves a mitogen-activated protein kinase pathway. The Journal of neuroscience : the official journal of the Society for Neuroscience, 22(12), 4860-8.
[PubMed:12077183]
[PMC]
[WorldCat]
↑Sampson, K.J., & Kass, R.S. (2010).
Molecular mechanisms of adrenergic stimulation in the heart. Heart rhythm, 7(8), 1151-3.
[PubMed:20156590]
[PMC]
[WorldCat]
[DOI]
↑Wahl-Schott, C., & Biel, M. (2009).
HCN channels: structure, cellular regulation and physiological function. Cellular and molecular life sciences : CMLS, 66(3), 470-94.
[PubMed:18953682]
[WorldCat]
[DOI]
↑ 27.027.1Sampson, K.J., & Kass, R.S. (2010).
Molecular mechanisms of adrenergic stimulation in the heart. Heart rhythm, 7(8), 1151-3.
[PubMed:20156590]
[PMC]
[WorldCat]
[DOI]
↑Gray, P.C., Scott, J.D., & Catterall, W.A. (1998).
Regulation of ion channels by cAMP-dependent protein kinase and A-kinase anchoring proteins. Current opinion in neurobiology, 8(3), 330-4.
[PubMed:9687361]
[WorldCat]
[DOI]
↑Kamm, K.E., & Stull, J.T. (2001).
Dedicated myosin light chain kinases with diverse cellular functions. The Journal of biological chemistry, 276(7), 4527-30.
[PubMed:11096123]
[WorldCat]
[DOI]
↑Conti, M.A., & Adelstein, R.S. (1981).
The relationship between calmodulin binding and phosphorylation of smooth muscle myosin kinase by the catalytic subunit of 3':5' cAMP-dependent protein kinase. The Journal of biological chemistry, 256(7), 3178-81.
[PubMed:6259152]
[WorldCat]
↑Starke, K. (2001).
Presynaptic autoreceptors in the third decade: focus on alpha2-adrenoceptors. Journal of neurochemistry, 78(4), 685-93.
[PubMed:11520889]
[WorldCat]
[DOI]
↑Moore, R.Y., & Bloom, F.E. (1978).
Central catecholamine neuron systems: anatomy and physiology of the dopamine systems. Annual review of neuroscience, 1, 129-69.
[PubMed:756202]
[WorldCat]
[DOI]
↑Moruzzi, G., & Magoun, H.W. (1949).
Brain stem reticular formation and activation of the EEG. Electroencephalography and clinical neurophysiology, 1(4), 455-73.
[PubMed:18421835]
[WorldCat]
↑Aston-Jones, G., & Bloom, F.E. (1981).
Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. The Journal of neuroscience : the official journal of the Society for Neuroscience, 1(8), 876-86.
[PubMed:7346592]
[WorldCat]
↑Foote, S.L., Aston-Jones, G., & Bloom, F.E. (1980).
Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proceedings of the National Academy of Sciences of the United States of America, 77(5), 3033-7.
[PubMed:6771765]
[PMC]
[WorldCat]
[DOI]
↑Foote, S.L., Freedman, R., & Oliver, A.P. (1975).
Effects of putative neurotransmitters on neuronal activity in monkey auditory cortex. Brain research, 86(2), 229-42.
[PubMed:234774]
[WorldCat]
[DOI]
↑Waterhouse, B.D., & Woodward, D.J. (1980).
Interaction of norepinephrine with cerebrocortical activity evoked by stimulation of somatosensory afferent pathways in the rat. Experimental neurology, 67(1), 11-34.
[PubMed:7349980]
[WorldCat]
[DOI]
↑Segal, M., & Bloom, F.E. (1976).
The action of norepinephrine in the rat hippocampus. IV. The effects of locus coeruleus stimulation on evoked hippocampal unit activity. Brain research, 107(3), 513-25.
[PubMed:178411]
[WorldCat]
[DOI]
↑Lecas, J.C. (2004).
Locus coeruleus activation shortens synaptic drive while decreasing spike latency and jitter in sensorimotor cortex. Implications for neuronal integration. The European journal of neuroscience, 19(9), 2519-30.
[PubMed:15128405]
[WorldCat]
[DOI]
↑Bouret, S., & Sara, S.J. (2002).
Locus coeruleus activation modulates firing rate and temporal organization of odour-induced single-cell responses in rat piriform cortex. The European journal of neuroscience, 16(12), 2371-82.
[PubMed:12492432]
[WorldCat]
[DOI]
↑Mair, R.D., Zhang, Y., Bailey, K.R., Toupin, M.M., & Mair, R.G. (2005).
Effects of clonidine in the locus coeruleus on prefrontal- and hippocampal-dependent measures of attention and memory in the rat. Psychopharmacology, 181(2), 280-8.
[PubMed:15830223]
[WorldCat]
[DOI]
↑Ramos, B.P., & Arnsten, A.F. (2007).
Adrenergic pharmacology and cognition: focus on the prefrontal cortex. Pharmacology & therapeutics, 113(3), 523-36.
[PubMed:17303246]
[PMC]
[WorldCat]
[DOI]
↑Wang, M., Ramos, B.P., Paspalas, C.D., Shu, Y., Simen, A., Duque, A., ..., & Arnsten, A.F. (2007).
Alpha2A-adrenoceptors strengthen working memory networks by inhibiting cAMP-HCN channel signaling in prefrontal cortex. Cell, 129(2), 397-410.
[PubMed:17448997]
[WorldCat]
[DOI]
↑Coull, J.T., Büchel, C., Friston, K.J., & Frith, C.D. (1999).
Noradrenergically mediated plasticity in a human attentional neuronal network. NeuroImage, 10(6), 705-15.
[PubMed:10600416]
[WorldCat]
[DOI]
↑McGaugh, J.L., & Roozendaal, B. (2009).
Drug enhancement of memory consolidation: historical perspective and neurobiological implications. Psychopharmacology, 202(1-3), 3-14.
[PubMed:18704369]
[WorldCat]
[DOI]
↑Sara, S.J., Roullet, P., & Przybyslawski, J. (1999).
Consolidation of memory for odor-reward association: beta-adrenergic receptor involvement in the late phase. Learning & memory (Cold Spring Harbor, N.Y.), 6(2), 88-96.
[PubMed:10327234]
[PMC]
[WorldCat]
↑Tronel, S., Feenstra, M.G., & Sara, S.J. (2004).
Noradrenergic action in prefrontal cortex in the late stage of memory consolidation. Learning & memory (Cold Spring Harbor, N.Y.), 11(4), 453-8.
[PubMed:15254217]
[PMC]
[WorldCat]
[DOI]
↑Sara, S.J., & Devauges, V. (1988).
Priming stimulation of locus coeruleus facilitates memory retrieval in the rat. Brain research, 438(1-2), 299-303.
[PubMed:3345434]
[WorldCat]
[DOI]
↑Sara, S.J., & Devauges, V. (1989).
Idazoxan, an alpha-2 antagonist, facilitates memory retrieval in the rat. Behavioral and neural biology, 51(3), 401-11.
[PubMed:2543356]
[WorldCat]
[DOI]
↑Murchison, C.F., Zhang, X.Y., Zhang, W.P., Ouyang, M., Lee, A., & Thomas, S.A. (2004).
A distinct role for norepinephrine in memory retrieval. Cell, 117(1), 131-43.
[PubMed:15066288]
[WorldCat]
[DOI]
↑ 52.052.1Strawn, J.R., & Geracioti, T.D. (2008).
Noradrenergic dysfunction and the psychopharmacology of posttraumatic stress disorder. Depression and anxiety, 25(3), 260-71.
[PubMed:17354267]
[WorldCat]
[DOI]
↑Debiec, J., & LeDoux, J.E. (2006).
Noradrenergic signaling in the amygdala contributes to the reconsolidation of fear memory: treatment implications for PTSD. Annals of the New York Academy of Sciences, 1071, 521-4.
[PubMed:16891611]
[WorldCat]
[DOI]
↑Diergaarde, L., Schoffelmeer, A.N., & De Vries, T.J. (2008).
Pharmacological manipulation of memory reconsolidation: towards a novel treatment of pathogenic memories. European journal of pharmacology, 585(2-3), 453-7.
[PubMed:18410917]
[WorldCat]
[DOI]
↑Brunet, A., Orr, S.P., Tremblay, J., Robertson, K., Nader, K., & Pitman, R.K. (2008).
Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumatic stress disorder. Journal of psychiatric research, 42(6), 503-6.
[PubMed:17588604]
[WorldCat]
[DOI]
↑E R Kandel, J H Schwartz, T M Jessell Principles of Neural Science, Fourth Edition Mc Graw Hill (New York):2000
↑N R Carlson Physiology of Behavior, Tenth Edition Pearson Education (Boston):2009