「アミロイドーシス」の版間の差分
細編集の要約なし |
細編集の要約なし |
||
| (4人の利用者による、間の21版が非表示) | |||
| 2行目: | 2行目: | ||
<font size="+1">[http://researchmap.jp/_tomitataisuke 富田 泰輔]</font><br> | <font size="+1">[http://researchmap.jp/_tomitataisuke 富田 泰輔]</font><br> | ||
''東京大学 薬学研究科''<br> | ''東京大学 薬学研究科''<br> | ||
DOI [[XXXX]]/XXXX 原稿受付日:2013年11月27日 原稿完成日:2013年月日<br> | |||
担当編集委員:[http://researchmap.jp/read0141446 漆谷 真] | 担当編集委員:[http://researchmap.jp/read0141446 漆谷 真](京都大学 大学院医学研究科)<br> | ||
</div> | </div> | ||
英:amyloidosis | |||
{{box|text= [[アミロイド]]amyloidは[[wikipedia:ja:コンゴーレッド|コンゴーレッド]]染色でオレンジ色に染まり、[[wikipedia:ja:偏光顕微鏡|偏光顕微鏡]]で緑色偏光を呈し、[[wikipedia:ja:電子顕微鏡|電子顕微鏡]] | {{box|text= [[アミロイド]]amyloidは[[wikipedia:ja:コンゴーレッド|コンゴーレッド]]染色でオレンジ色に染まり、[[wikipedia:ja:偏光顕微鏡|偏光顕微鏡]]で緑色偏光を呈し、[[wikipedia:ja:電子顕微鏡|電子顕微鏡]]観察下では7~15nmの繊維構造を呈する物質として定義される。アミロイドが、組織間隙に沈着して臓器の機能不全が生じる疾患をアミロイドーシス amyloidosisと呼ぶ<ref><pubmed> 22664198 </pubmed></ref>。アミロイドタンパク質の種類や臓器によって特徴が見られ、大きく[[全身性アミロイドーシス]]と[[限局性アミロイドーシス]]に分類される。代表的な全身性アミロイドーシスには、全身性AAアミロイドーシス、家族性アミロイドニューロパチーが挙げられる。限局性アミロイドーシスには脳アミロイドーシスである、[[アルツハイマー病]]、[[脳血管アミロイドアンギオパチー]]、遺伝性アミロイド性脳出血で、[[クロイツフェルト・ヤコブ病]]などが知られている。}} | ||
(編集コメント:元の抄録はアミロイドそのものの抄録でしたので、アミロイドーシスの内容としました) | |||
==アミロイドーシスとは== | ==アミロイドーシスとは== | ||
アミロイドamyloidはコンゴーレッド染色でオレンジ色に染まり、偏光顕微鏡で緑色偏光を呈し、電子顕微鏡観察下では7~15nmの繊維構造を呈する物質として定義される。アミロイドが、組織間隙に沈着する疾患を総称してアミロイドーシス amyloidosisと呼ぶ<ref><pubmed> 22664198 </pubmed></ref>。多くの場合、前駆タンパクであるアミロイドタンパク質が折りたたみ障害を引き起こして重合し、[[wikipedia:ja:βシート|βシート]]構造に富む不溶性線維として蓄積・凝集している。基本的には、アミロイドーシス発症の分子病態は凝集するアミロイドタンパク質の濃度上昇か、凝集能亢進によるものである。したがってアミロイドタンパク質の除去が根本治療戦略となる。 | |||
沈着するアミロイドタンパク質の種類や臓器によって特徴が見られ、特に大きく全身性アミロイドーシスと限局性アミロイドーシスに分類されている。 | |||
(編集コメント:病名、原因物質と沈着するタンパク質を表か箇条書きに出来ないでしょうか。) | |||
== 分類 == | |||
===全身性アミロイドーシス=== | ===全身性アミロイドーシス=== | ||
アミロイドタンパク質が血中に存在する場合は全身性アミロイドーシスとなる<ref><pubmed> 23451869 </pubmed></ref>。 | アミロイドタンパク質が血中に存在する場合は全身性アミロイドーシスとなる<ref><pubmed> 23451869 </pubmed></ref>。 | ||
| 22行目: | 25行目: | ||
アミロイドタンパク質としては、モノクローナル[[免疫グロブリン]]の[[wikipedia:ja:L鎖|L鎖]]由来の[[アミロイドAL]]や[[wikipedia:ja:H鎖|H鎖]]由来の[[アミロイドAH]]、[[wikipedia:ja:血清アミロイドA|血清アミロイドA]]の代謝産物である[[アミロイドA]](AA)、β2[[ミクログロブリン]]、[[トランスサイレチン]]、[[ゲルソリン]]、[[アポAI]]が知られている。いずれもアミロイドタンパク質の産生亢進、濃度上昇がアミロイドーシスを惹起していることが知られており、例えばアミロイドALでは免疫グロブリン産生細胞である[[wikipedia:ja:形質細胞|形質細胞]]の過剰な増殖や腫瘍化がその原因である。また[[wikipedia:ja:膠原病|膠原病]]や[[wikipedia:ja:リウマチ|リウマチ]]などが原因となり全身性[[wikipedia:ja:慢性炎症|慢性炎症]]を基礎疾患として血清アミロイドAの濃度上昇が継続し、全身性AAアミロイドーシスを惹起する。さらに[[wikipedia:ja:腎障害|腎障害]]及び[[wikipedia:ja:血液透析|血液透析]]によってβ2ミクログロブリンの排泄、除去が不全となり、10年以上の長期透析の結果アミロイド沈着を招くことが知られている。 | アミロイドタンパク質としては、モノクローナル[[免疫グロブリン]]の[[wikipedia:ja:L鎖|L鎖]]由来の[[アミロイドAL]]や[[wikipedia:ja:H鎖|H鎖]]由来の[[アミロイドAH]]、[[wikipedia:ja:血清アミロイドA|血清アミロイドA]]の代謝産物である[[アミロイドA]](AA)、β2[[ミクログロブリン]]、[[トランスサイレチン]]、[[ゲルソリン]]、[[アポAI]]が知られている。いずれもアミロイドタンパク質の産生亢進、濃度上昇がアミロイドーシスを惹起していることが知られており、例えばアミロイドALでは免疫グロブリン産生細胞である[[wikipedia:ja:形質細胞|形質細胞]]の過剰な増殖や腫瘍化がその原因である。また[[wikipedia:ja:膠原病|膠原病]]や[[wikipedia:ja:リウマチ|リウマチ]]などが原因となり全身性[[wikipedia:ja:慢性炎症|慢性炎症]]を基礎疾患として血清アミロイドAの濃度上昇が継続し、全身性AAアミロイドーシスを惹起する。さらに[[wikipedia:ja:腎障害|腎障害]]及び[[wikipedia:ja:血液透析|血液透析]]によってβ2ミクログロブリンの排泄、除去が不全となり、10年以上の長期透析の結果アミロイド沈着を招くことが知られている。 | ||
遺伝子変異によって生じる全身性アミロイドーシスとして、[[wikipedia:ja:家族性アミロイドニューロパチー|家族性アミロイドニューロパチー]] [[wikipedia:Familial amyloid polyneuropathy|Familial amyloid polyneuropathy(FAP)]]が知られている<ref><pubmed> 22094129 </pubmed></ref>。FAPはトランスサイレチン、ゲルソリン、アポAI、血清アミロイドA遺伝子変異に連鎖し、これらのアミロイドタンパク質が[[神経節]]を含む神経系および他の臓器に沈着する。また最近になり、全身性アミロイドーシスを惹起する[[プリオン]] | 遺伝子変異によって生じる全身性アミロイドーシスとして、[[wikipedia:ja:家族性アミロイドニューロパチー|家族性アミロイドニューロパチー]] [[wikipedia:Familial amyloid polyneuropathy|Familial amyloid polyneuropathy(FAP)]]が知られている<ref><pubmed> 22094129 </pubmed></ref>。FAPはトランスサイレチン、ゲルソリン、アポAI、血清アミロイドA遺伝子変異に連鎖し、これらのアミロイドタンパク質が[[神経節]]を含む神経系および他の臓器に沈着する。また最近になり、全身性アミロイドーシスを惹起する[[プリオン]]遺伝子も同定された<ref><pubmed> 24224623 </pubmed></ref>。我が国を含めて、特にトランスサイレチン遺伝子変異によるFAPが最も多い<ref><pubmed> 11940682 </pubmed></ref>。 | ||
通常トランスサイレチンは四量体を形成しているが、遺伝子変異によって生じるアミノ酸置換によって不安定な単量体へ解離しやすくなり、なんらかの機序で重合して線維化すると考えられている。体内のトランスサイレチンは主として肝臓で産生されるが、肝実質にアミロイドは沈着しない。このためFAP患者の肝臓を移植により正常肝に換えることでアミロイドタンパク質である変異トランスサイレチンの消失が期待され、移植後多くの症例でFAPの臨床進行が停止するか、遅延することが確認されている。また2013年には、トランスサイレチンの四量体の解離及び変性を抑制することでアミロイド形成を阻害し、[[末梢神経]]障害の進行を抑制する[[wikipedia:Vyndaqel|Vyndaqel]](一般名:[[wikipedia:en:Tafamidis|Tafamidis]])が承認された。 | 通常トランスサイレチンは四量体を形成しているが、遺伝子変異によって生じるアミノ酸置換によって不安定な単量体へ解離しやすくなり、なんらかの機序で重合して線維化すると考えられている。体内のトランスサイレチンは主として肝臓で産生されるが、肝実質にアミロイドは沈着しない。このためFAP患者の肝臓を移植により正常肝に換えることでアミロイドタンパク質である変異トランスサイレチンの消失が期待され、移植後多くの症例でFAPの臨床進行が停止するか、遅延することが確認されている。また2013年には、トランスサイレチンの四量体の解離及び変性を抑制することでアミロイド形成を阻害し、[[末梢神経]]障害の進行を抑制する[[wikipedia:Vyndaqel|Vyndaqel]](一般名:[[wikipedia:en:Tafamidis|Tafamidis]])が承認された。 | ||
===限局性アミロイドーシス=== | ===限局性アミロイドーシス=== | ||
| 35行目: | 36行目: | ||
その他の限局性アミロイドーシスとしては、内分泌アミロイドーシスのアミロイドタンパク質としては[[カルシトニン]]、[[アミリン]]、[[インスリン]]、[[心房ナトリウム利尿ペプチド]]が同定されており、主にこれらのホルモンを分泌する細胞由来の腫瘍内で蓄積・沈着が観察される。また皮膚アミロイドーシスとしては[[wikipedia:ja:ケラチン|ケラチン]]が、限局性結節性アミロイドーシスはアミロイドALがアミロイドタンパク質として蓄積することが報告されている。 | その他の限局性アミロイドーシスとしては、内分泌アミロイドーシスのアミロイドタンパク質としては[[カルシトニン]]、[[アミリン]]、[[インスリン]]、[[心房ナトリウム利尿ペプチド]]が同定されており、主にこれらのホルモンを分泌する細胞由来の腫瘍内で蓄積・沈着が観察される。また皮膚アミロイドーシスとしては[[wikipedia:ja:ケラチン|ケラチン]]が、限局性結節性アミロイドーシスはアミロイドALがアミロイドタンパク質として蓄積することが報告されている。 | ||
(編集コメント:通常疾患を扱う項目は、臨床症状、検査所見、診断(鑑別診断)、病態生理、治療、疫学(発症率など)の順で書いて頂くようにしております。種々の疾患の総称なので難しいかと思いますが、なるべくこれに沿ってイ記述して頂ければと思います。漆谷先生、最終判断は御任せ致します。) | |||
==病態生理== | ==病態生理== | ||
[[Image:2M5N.pdb|thumb|350px|''' | [[Image:2M5N.pdb|thumb|350px|'''図1.クロスβ構造'''<br>トランスサイレチン部分ペプチドからなるクロスβ構造。PDB ID: {{PDB2|2M5N}}]] | ||
(編集コメント:結晶構造は回転できる物に取り替えました。これで良いかご確認下さい。SafariではWebGLをonにして下さい。) | |||
===構造=== | ===構造=== | ||
各アミロイドタンパク質には一定の共通したアミノ酸配列や構造は見られないが、アミロイド線維になると共通して[[クロスβ構造]]と呼ばれる形態をとっている<ref><pubmed> 17468747 </pubmed></ref><ref><pubmed> 21456964 </pubmed></ref><ref><pubmed> 23513222 </pubmed></ref>。これはアミロイド線維を構成するポリペプチド鎖が線維軸と垂直方向に[[wikipedia:ja:βストランド|βストランド]]となり、かつ線維軸方向に[[wikipedia:ja:βシート構造|βシート構造]] | 各アミロイドタンパク質には一定の共通したアミノ酸配列や構造は見られないが、アミロイド線維になると共通して[[クロスβ構造]]と呼ばれる形態をとっている<ref><pubmed> 17468747 </pubmed></ref><ref><pubmed> 21456964 </pubmed></ref><ref><pubmed> 23513222 </pubmed></ref>。これはアミロイド線維を構成するポリペプチド鎖が線維軸と垂直方向に[[wikipedia:ja:βストランド|βストランド]]となり、かつ線維軸方向に[[wikipedia:ja:βシート構造|βシート構造]]をとっているものである。このような構造学的特徴はイメージング技術に応用されつつあり、[[wikipedia:ja:Aβ|Aβ]]線維に特異的に結合する低分子化合物を利用した[[アミロイドPETスキャン]]が可能となった<ref><pubmed> 14991808 </pubmed></ref><ref><pubmed> 21245183 </pubmed></ref>。 | ||
===線維形成過程と伝播=== | ===線維形成過程と伝播=== | ||
[[Image:2M4J.pdb|thumb|350px|'''図3.アルツハイマー病患者脳由来のAβ線維構造'''<br>患者脳由来アミロイドから伸長した[[アミロイドβタンパク質]]の分子構造。PDB ID: {{PDB2|2M4J}}]] | |||
[[Image:2M4J.pdb|thumb|350px|''' | |||
アミロイド線維形成過程では、多くの場合正常なフォールディングをうけているアミロイドタンパク質が何らかの理由で一旦部分変性し、会合することが必要である。また線維形成過程はその鋳型となるシード(種、核)の形成を契機として急速に進んでいくことが示されている<ref><pubmed> 22885025 </pubmed></ref> | (編集コメント:長かったので、小見出しをつけました。内容に照らして適当かご確認下さい。) | ||
アミロイド線維形成過程では、多くの場合正常なフォールディングをうけているアミロイドタンパク質が何らかの理由で一旦部分変性し、会合することが必要である。また線維形成過程はその鋳型となるシード(種、核)の形成を契機として急速に進んでいくことが示されている<ref><pubmed> 22885025 </pubmed></ref>。すなわち、このシードの両端の末端にアミロイドタンパク質が結合して線維が伸長していくと考えられている。 | |||
このようなシード依存性伸長反応モデルは、[[プリオン]]タンパク質が示す伝播能力とも関連していると考えられている。すなわち、一旦異常構造をとったタンパク質がシードとなり、別の個体におけるアミロイドタンパク質の構造及び性質を変化させていくというモデルである<ref><pubmed> 8513491 </pubmed></ref>。またシードへの組み込みはアミロイドタンパク質が同様の構造を取りうるかどうかに依存する。プリオンの感染性にはごく僅かなアミノ酸の違いに起因する「種の壁」が存在するが、この現象も一次配列の違いに依存する各種のプリオンが形成するシード構造の違いによって説明できる。 | このようなシード依存性伸長反応モデルは、[[プリオン]]タンパク質が示す伝播能力とも関連していると考えられている。すなわち、一旦異常構造をとったタンパク質がシードとなり、別の個体におけるアミロイドタンパク質の構造及び性質を変化させていくというモデルである<ref><pubmed> 8513491 </pubmed></ref>[[Image:TTfig6.png|thumb|350px|'''図2.アミロイド線維形成過程'''<br>アミロイド線維形成過程におけるシードの役割]]。またシードへの組み込みはアミロイドタンパク質が同様の構造を取りうるかどうかに依存する。プリオンの感染性にはごく僅かなアミノ酸の違いに起因する「種の壁」が存在するが、この現象も一次配列の違いに依存する各種のプリオンが形成するシード構造の違いによって説明できる。 | ||
最近ではアミロイドを形成しうるアミロイドタンパク質がいずれもプリオン様の伝播能力を示す可能性が推測されている<ref><pubmed> 22660329 </pubmed></ref><ref><pubmed> 24005412 </pubmed></ref>。実際、全身性アミロイドーシスの一つであるAAアミロイドーシスはモデル[[wikipedia:ja:マウス|マウス]]を用いた伝播実験が確認されているが、野生の[[wikipedia:ja:チーター|チーター]]においてAAアミロイドーシス発症頻度が近年上昇していることが示されていた。そして興味深いことに、AAアミロイドーシスを発症した個体の糞に伝播性が極めて高いアミロイドA線維が含まれていることが明らかとなった<ref><pubmed> 18474855 </pubmed></ref>。 | 最近ではアミロイドを形成しうるアミロイドタンパク質がいずれもプリオン様の伝播能力を示す可能性が推測されている<ref><pubmed> 22660329 </pubmed></ref><ref><pubmed> 24005412 </pubmed></ref>。実際、全身性アミロイドーシスの一つであるAAアミロイドーシスはモデル[[wikipedia:ja:マウス|マウス]]を用いた伝播実験が確認されているが、野生の[[wikipedia:ja:チーター|チーター]]においてAAアミロイドーシス発症頻度が近年上昇していることが示されていた。そして興味深いことに、AAアミロイドーシスを発症した個体の糞に伝播性が極めて高いアミロイドA線維が含まれていることが明らかとなった<ref><pubmed> 18474855 </pubmed></ref>。 | ||
糞便を介したアミロイドーシス伝播は、野生動物におけるプリオン病([[wikipedia:ja:ヒツジ|ヒツジ]]おける[[スクレイピー]]、[[wikipedia:ja:シカ|シカ]]における[[Chronic wasting disease | 糞便を介したアミロイドーシス伝播は、野生動物におけるプリオン病([[wikipedia:ja:ヒツジ|ヒツジ]]おける[[スクレイピー]]、[[wikipedia:ja:シカ|シカ]]における[[Chronic wasting disease[[)の[[wikipedia:ja:水平伝播|水平伝播]]メカニズムを説明できるものとして注目を集めている。特に末梢神経や[[wikipedia:ja:リンパ節|リンパ節]]を介したプリオンの伝播に関しては、食物摂取などを介した末梢組織から生じうる限局性アミロイドーシスの発症機構を担っている可能性がある<ref><pubmed> 24189576 </pubmed></ref>。またAβについても、アルツハイマー病モデルマウスの[[wikipedia:ja:腹腔|腹腔]]内にAβ線維を注入すると[[大脳皮質]]でのAβの沈着が亢進することも示されている<ref><pubmed> 20966215 </pubmed></ref>。 | ||
このようなタンパク質凝集物の細胞間伝播という概念は必ずしもアミロイドの形成には依存しておらず、凝集して線維を形成するタンパク質に普遍的に観察される可能性があり、最近では様々な神経変性疾患において細胞内に蓄積するタンパク質([[タウ]]、[[シヌクレイン]]、[[TDP-43]]など)においても伝播能力の存在が確認されつつある<ref><pubmed> 24005412 </pubmed></ref>。また[[wikipedia:ja:酵母|酵母]]などにおいてはプリオン様タンパク性因子による形質転換が報告されており、タンパク質の構造変化に依存した形質の伝播様式として注目されている<ref><pubmed> 23379365 </pubmed></ref>。 | このようなタンパク質凝集物の細胞間伝播という概念は必ずしもアミロイドの形成には依存しておらず、凝集して線維を形成するタンパク質に普遍的に観察される可能性があり、最近では様々な神経変性疾患において細胞内に蓄積するタンパク質([[タウ]]、[[シヌクレイン]]、[[TDP-43]]など)においても伝播能力の存在が確認されつつある<ref><pubmed> 24005412 </pubmed></ref>。また[[wikipedia:ja:酵母|酵母]]などにおいてはプリオン様タンパク性因子による形質転換が報告されており、タンパク質の構造変化に依存した形質の伝播様式として注目されている<ref><pubmed> 23379365 </pubmed></ref>。 | ||
一方、アルツハイマー病患者脳から得られたAβ線維の構造解析がなされ、<i>in vitro</i>で凝集させた構造とは異なる凝集形態を示していたことから、<i>in vivo</i>における凝集プロセスの違いが指摘されており<ref><pubmed> 24034249 </pubmed></ref>、伝播メカニズムとの関係の解明が待たれている。 | |||
===細胞毒性=== | ===細胞毒性=== | ||
アミロイド線維が発揮する細胞障害および毒性はアミロイドーシスにおける臓器不全の基本的病態と言える。アミロイド沈着後に生じる疾患プロセスを抑制する治療薬の開発のためにも、その理解は必須である。しかしアミロイドタンパク質のどのような構造、分子状態が毒性を発揮するのかについては未だ明確ではない。近年ではAβとFAD変異がもたらす分子病態の解析から、アミロイド線維そのものではなく、その中間体となるオリゴマー<ref><pubmed> 12702875 </pubmed></ref>に起因しているというオリゴマー仮説が提唱されている。 | アミロイド線維が発揮する細胞障害および毒性はアミロイドーシスにおける臓器不全の基本的病態と言える。アミロイド沈着後に生じる疾患プロセスを抑制する治療薬の開発のためにも、その理解は必須である。しかしアミロイドタンパク質のどのような構造、分子状態が毒性を発揮するのかについては未だ明確ではない。近年ではAβとFAD変異がもたらす分子病態の解析から、アミロイド線維そのものではなく、その中間体となるオリゴマー<ref><pubmed> 12702875 </pubmed></ref>に起因しているというオリゴマー仮説が提唱されている。 | ||
このアミロイドタンパク質の凝集物がどのように細胞傷害を惹起しているか、という点については、[[脂質二重膜]]の障害、[[酸化的ストレス]]や[[小胞体ストレス]]の惹起、[[ミトコンドリア]]障害などが想定されている<ref><pubmed> 23820032 </pubmed></ref>。興味深いことに、全く異なるアミロイド原性タンパク質であるAβとADanが脳実質に蓄積するそれぞれの疾患モデルマウスを、神経障害と関連するtauトランスジェニックマウスと交配すると、いずれの場合もtau病理が亢進されることが示された<ref><pubmed> 20385796 </pubmed></ref>。これは少なくとも[[大脳皮質]] | このアミロイドタンパク質の凝集物がどのように細胞傷害を惹起しているか、という点については、[[脂質二重膜]]の障害、[[酸化的ストレス]]や[[小胞体ストレス]]の惹起、[[ミトコンドリア]]障害などが想定されている<ref><pubmed> 23820032 </pubmed></ref>。興味深いことに、全く異なるアミロイド原性タンパク質であるAβとADanが脳実質に蓄積するそれぞれの疾患モデルマウスを、神経障害と関連するtauトランスジェニックマウスと交配すると、いずれの場合もtau病理が亢進されることが示された<ref><pubmed> 20385796 </pubmed></ref>。これは少なくとも[[大脳皮質]]に沈着するアミロイドが示す神経細胞傷害プロセスの下流には共通性があることを示唆している。すなわち、アミロイド原性タンパク質の種類を問わず、どのような線維がどの細胞や臓器に沈着するかによって最終的にアミロイドーシスにおける病態が決定する可能性が考えられている。またAβが細胞外から神経細胞毒性を呈するために毒性受容体が想定さられており、[[NMDA型]]および[[AMPA型グルタミン酸]]受容体、α7[[ニコチン性アセチルコリン受容体]]、[[インスリン受容体]]、[[RAGE]]、プリオンタンパク質や[[EPHB2|EphB2]]、LilrB2などがその候補として挙げられている。 | ||
==関連項目== | ==関連項目== | ||
| 176行目: | 72行目: | ||
*[[プリオン病]] | *[[プリオン病]] | ||
*[[アミロイドβタンパク質]] | *[[アミロイドβタンパク質]] | ||
==参考文献== | ===参考文献=== | ||
<references/> | <references/> | ||
2013年12月9日 (月) 11:33時点における版
英:amyloidosis
アミロイドamyloidはコンゴーレッド染色でオレンジ色に染まり、偏光顕微鏡で緑色偏光を呈し、電子顕微鏡観察下では7~15nmの繊維構造を呈する物質として定義される。アミロイドが、組織間隙に沈着して臓器の機能不全が生じる疾患をアミロイドーシス amyloidosisと呼ぶ[1]。アミロイドタンパク質の種類や臓器によって特徴が見られ、大きく全身性アミロイドーシスと限局性アミロイドーシスに分類される。代表的な全身性アミロイドーシスには、全身性AAアミロイドーシス、家族性アミロイドニューロパチーが挙げられる。限局性アミロイドーシスには脳アミロイドーシスである、アルツハイマー病、脳血管アミロイドアンギオパチー、遺伝性アミロイド性脳出血で、クロイツフェルト・ヤコブ病などが知られている。
(編集コメント:元の抄録はアミロイドそのものの抄録でしたので、アミロイドーシスの内容としました)
アミロイドーシスとは
アミロイドamyloidはコンゴーレッド染色でオレンジ色に染まり、偏光顕微鏡で緑色偏光を呈し、電子顕微鏡観察下では7~15nmの繊維構造を呈する物質として定義される。アミロイドが、組織間隙に沈着する疾患を総称してアミロイドーシス amyloidosisと呼ぶ[2]。多くの場合、前駆タンパクであるアミロイドタンパク質が折りたたみ障害を引き起こして重合し、βシート構造に富む不溶性線維として蓄積・凝集している。基本的には、アミロイドーシス発症の分子病態は凝集するアミロイドタンパク質の濃度上昇か、凝集能亢進によるものである。したがってアミロイドタンパク質の除去が根本治療戦略となる。
沈着するアミロイドタンパク質の種類や臓器によって特徴が見られ、特に大きく全身性アミロイドーシスと限局性アミロイドーシスに分類されている。
(編集コメント:病名、原因物質と沈着するタンパク質を表か箇条書きに出来ないでしょうか。)
分類
全身性アミロイドーシス
アミロイドタンパク質が血中に存在する場合は全身性アミロイドーシスとなる[3]。
アミロイドタンパク質としては、モノクローナル免疫グロブリンのL鎖由来のアミロイドALやH鎖由来のアミロイドAH、血清アミロイドAの代謝産物であるアミロイドA(AA)、β2ミクログロブリン、トランスサイレチン、ゲルソリン、アポAIが知られている。いずれもアミロイドタンパク質の産生亢進、濃度上昇がアミロイドーシスを惹起していることが知られており、例えばアミロイドALでは免疫グロブリン産生細胞である形質細胞の過剰な増殖や腫瘍化がその原因である。また膠原病やリウマチなどが原因となり全身性慢性炎症を基礎疾患として血清アミロイドAの濃度上昇が継続し、全身性AAアミロイドーシスを惹起する。さらに腎障害及び血液透析によってβ2ミクログロブリンの排泄、除去が不全となり、10年以上の長期透析の結果アミロイド沈着を招くことが知られている。
遺伝子変異によって生じる全身性アミロイドーシスとして、家族性アミロイドニューロパチー Familial amyloid polyneuropathy(FAP)が知られている[4]。FAPはトランスサイレチン、ゲルソリン、アポAI、血清アミロイドA遺伝子変異に連鎖し、これらのアミロイドタンパク質が神経節を含む神経系および他の臓器に沈着する。また最近になり、全身性アミロイドーシスを惹起するプリオン遺伝子も同定された[5]。我が国を含めて、特にトランスサイレチン遺伝子変異によるFAPが最も多い[6]。
通常トランスサイレチンは四量体を形成しているが、遺伝子変異によって生じるアミノ酸置換によって不安定な単量体へ解離しやすくなり、なんらかの機序で重合して線維化すると考えられている。体内のトランスサイレチンは主として肝臓で産生されるが、肝実質にアミロイドは沈着しない。このためFAP患者の肝臓を移植により正常肝に換えることでアミロイドタンパク質である変異トランスサイレチンの消失が期待され、移植後多くの症例でFAPの臨床進行が停止するか、遅延することが確認されている。また2013年には、トランスサイレチンの四量体の解離及び変性を抑制することでアミロイド形成を阻害し、末梢神経障害の進行を抑制するVyndaqel(一般名:Tafamidis)が承認された。
限局性アミロイドーシス
特定の臓器に限局して沈着を認める場合は限局性アミロイドーシスとなる。臓器に応じて分類され、脳アミロイドーシス[7]としてはアルツハイマー病や脳血管アミロイドアンギオパチーで蓄積が見られるアミロイドβタンパク質(Aβ)の他、シスタチンCの遺伝子変異[8]がアイスランド型遺伝性アミロイド性脳出血で見出されている。
またプリオンタンパク質の蓄積、沈着はクロイツフェルト・ヤコブ病やゲルストマン・ストロイスラー・シャインカー症候群などのプリオン病患者脳で報告されている。さらにBRI2遺伝子の変異によって生じるアミロイドペプチドABri、ADanはそれぞれBritish型、Danish型家族性認知症患者脳において蓄積している[9]。BRI2はその最C末端部がFurinによって切断され分泌されているが、野生型ペプチドには凝集性が認められない。しかし終止コドン近傍の遺伝子変異により野生型よりも僅かに長く、凝集性の高いペプチドが分泌され、これらがアミロイドとして脳実質に蓄積する。
その他の限局性アミロイドーシスとしては、内分泌アミロイドーシスのアミロイドタンパク質としてはカルシトニン、アミリン、インスリン、心房ナトリウム利尿ペプチドが同定されており、主にこれらのホルモンを分泌する細胞由来の腫瘍内で蓄積・沈着が観察される。また皮膚アミロイドーシスとしてはケラチンが、限局性結節性アミロイドーシスはアミロイドALがアミロイドタンパク質として蓄積することが報告されている。
(編集コメント:通常疾患を扱う項目は、臨床症状、検査所見、診断(鑑別診断)、病態生理、治療、疫学(発症率など)の順で書いて頂くようにしております。種々の疾患の総称なので難しいかと思いますが、なるべくこれに沿ってイ記述して頂ければと思います。漆谷先生、最終判断は御任せ致します。)
病態生理
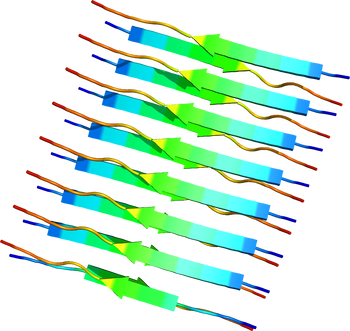
トランスサイレチン部分ペプチドからなるクロスβ構造。PDB ID: 2M5N
(編集コメント:結晶構造は回転できる物に取り替えました。これで良いかご確認下さい。SafariではWebGLをonにして下さい。)
構造
各アミロイドタンパク質には一定の共通したアミノ酸配列や構造は見られないが、アミロイド線維になると共通してクロスβ構造と呼ばれる形態をとっている[10][11][12]。これはアミロイド線維を構成するポリペプチド鎖が線維軸と垂直方向にβストランドとなり、かつ線維軸方向にβシート構造をとっているものである。このような構造学的特徴はイメージング技術に応用されつつあり、Aβ線維に特異的に結合する低分子化合物を利用したアミロイドPETスキャンが可能となった[13][14]。
線維形成過程と伝播

患者脳由来アミロイドから伸長したアミロイドβタンパク質の分子構造。PDB ID: 2M4J
(編集コメント:長かったので、小見出しをつけました。内容に照らして適当かご確認下さい。)
アミロイド線維形成過程では、多くの場合正常なフォールディングをうけているアミロイドタンパク質が何らかの理由で一旦部分変性し、会合することが必要である。また線維形成過程はその鋳型となるシード(種、核)の形成を契機として急速に進んでいくことが示されている[15]。すなわち、このシードの両端の末端にアミロイドタンパク質が結合して線維が伸長していくと考えられている。
このようなシード依存性伸長反応モデルは、プリオンタンパク質が示す伝播能力とも関連していると考えられている。すなわち、一旦異常構造をとったタンパク質がシードとなり、別の個体におけるアミロイドタンパク質の構造及び性質を変化させていくというモデルである[16]

アミロイド線維形成過程におけるシードの役割
。またシードへの組み込みはアミロイドタンパク質が同様の構造を取りうるかどうかに依存する。プリオンの感染性にはごく僅かなアミノ酸の違いに起因する「種の壁」が存在するが、この現象も一次配列の違いに依存する各種のプリオンが形成するシード構造の違いによって説明できる。
最近ではアミロイドを形成しうるアミロイドタンパク質がいずれもプリオン様の伝播能力を示す可能性が推測されている[17][18]。実際、全身性アミロイドーシスの一つであるAAアミロイドーシスはモデルマウスを用いた伝播実験が確認されているが、野生のチーターにおいてAAアミロイドーシス発症頻度が近年上昇していることが示されていた。そして興味深いことに、AAアミロイドーシスを発症した個体の糞に伝播性が極めて高いアミロイドA線維が含まれていることが明らかとなった[19]。
糞便を介したアミロイドーシス伝播は、野生動物におけるプリオン病(ヒツジおけるスクレイピー、シカにおける[[Chronic wasting disease[[)の水平伝播メカニズムを説明できるものとして注目を集めている。特に末梢神経やリンパ節を介したプリオンの伝播に関しては、食物摂取などを介した末梢組織から生じうる限局性アミロイドーシスの発症機構を担っている可能性がある[20]。またAβについても、アルツハイマー病モデルマウスの腹腔内にAβ線維を注入すると大脳皮質でのAβの沈着が亢進することも示されている[21]。
このようなタンパク質凝集物の細胞間伝播という概念は必ずしもアミロイドの形成には依存しておらず、凝集して線維を形成するタンパク質に普遍的に観察される可能性があり、最近では様々な神経変性疾患において細胞内に蓄積するタンパク質(タウ、シヌクレイン、TDP-43など)においても伝播能力の存在が確認されつつある[22]。また酵母などにおいてはプリオン様タンパク性因子による形質転換が報告されており、タンパク質の構造変化に依存した形質の伝播様式として注目されている[23]。
一方、アルツハイマー病患者脳から得られたAβ線維の構造解析がなされ、in vitroで凝集させた構造とは異なる凝集形態を示していたことから、in vivoにおける凝集プロセスの違いが指摘されており[24]、伝播メカニズムとの関係の解明が待たれている。
細胞毒性
アミロイド線維が発揮する細胞障害および毒性はアミロイドーシスにおける臓器不全の基本的病態と言える。アミロイド沈着後に生じる疾患プロセスを抑制する治療薬の開発のためにも、その理解は必須である。しかしアミロイドタンパク質のどのような構造、分子状態が毒性を発揮するのかについては未だ明確ではない。近年ではAβとFAD変異がもたらす分子病態の解析から、アミロイド線維そのものではなく、その中間体となるオリゴマー[25]に起因しているというオリゴマー仮説が提唱されている。
このアミロイドタンパク質の凝集物がどのように細胞傷害を惹起しているか、という点については、脂質二重膜の障害、酸化的ストレスや小胞体ストレスの惹起、ミトコンドリア障害などが想定されている[26]。興味深いことに、全く異なるアミロイド原性タンパク質であるAβとADanが脳実質に蓄積するそれぞれの疾患モデルマウスを、神経障害と関連するtauトランスジェニックマウスと交配すると、いずれの場合もtau病理が亢進されることが示された[27]。これは少なくとも大脳皮質に沈着するアミロイドが示す神経細胞傷害プロセスの下流には共通性があることを示唆している。すなわち、アミロイド原性タンパク質の種類を問わず、どのような線維がどの細胞や臓器に沈着するかによって最終的にアミロイドーシスにおける病態が決定する可能性が考えられている。またAβが細胞外から神経細胞毒性を呈するために毒性受容体が想定さられており、NMDA型およびAMPA型グルタミン酸受容体、α7ニコチン性アセチルコリン受容体、インスリン受容体、RAGE、プリオンタンパク質やEphB2、LilrB2などがその候補として挙げられている。
関連項目
参考文献
- ↑
Radford, S.E., & Weissman, J.S. (2012).
Special issue: the molecular and cellular mechanisms of amyloidosis. Journal of molecular biology, 421(2-3), 139-41. [PubMed:22664198] [WorldCat] [DOI] - ↑
Radford, S.E., & Weissman, J.S. (2012).
Special issue: the molecular and cellular mechanisms of amyloidosis. Journal of molecular biology, 421(2-3), 139-41. [PubMed:22664198] [WorldCat] [DOI] - ↑
Blancas-Mejía, L.M., & Ramirez-Alvarado, M. (2013).
Systemic amyloidoses. Annual review of biochemistry, 82, 745-74. [PubMed:23451869] [PMC] [WorldCat] [DOI] - ↑
Planté-Bordeneuve, V., & Said, G. (2011).
Familial amyloid polyneuropathy. The Lancet. Neurology, 10(12), 1086-97. [PubMed:22094129] [WorldCat] [DOI] - ↑
Mead, S., Gandhi, S., Beck, J., Caine, D., Gallujipali, D., Carswell, C., ..., & Collinge, J. (2013).
A novel prion disease associated with diarrhea and autonomic neuropathy. The New England journal of medicine, 369(20), 1904-14. [PubMed:24224623] [PMC] [WorldCat] [DOI] - ↑
Ikeda, S., Nakazato, M., Ando, Y., & Sobue, G. (2002).
Familial transthyretin-type amyloid polyneuropathy in Japan: clinical and genetic heterogeneity. Neurology, 58(7), 1001-7. [PubMed:11940682] [WorldCat] [DOI] - ↑
Yamada, M., & Naiki, H. (2012).
Cerebral amyloid angiopathy. Progress in molecular biology and translational science, 107, 41-78. [PubMed:22482447] [WorldCat] [DOI] - ↑
Palsdottir, A., Abrahamson, M., Thorsteinsson, L., Arnason, A., Olafsson, I., Grubb, A., & Jensson, O. (1988).
Mutation in cystatin C gene causes hereditary brain haemorrhage. Lancet (London, England), 2(8611), 603-4. [PubMed:2900981] [WorldCat] [DOI] - ↑
Tsachaki, M., Ghiso, J., & Efthimiopoulos, S. (2008).
BRI2 as a central protein involved in neurodegeneration. Biotechnology journal, 3(12), 1548-54. [PubMed:19072909] [WorldCat] [DOI] - ↑
Sawaya, M.R., Sambashivan, S., Nelson, R., Ivanova, M.I., Sievers, S.A., Apostol, M.I., ..., & Eisenberg, D. (2007).
Atomic structures of amyloid cross-beta spines reveal varied steric zippers. Nature, 447(7143), 453-7. [PubMed:17468747] [WorldCat] [DOI] - ↑
Toyama, B.H., & Weissman, J.S. (2011).
Amyloid structure: conformational diversity and consequences. Annual review of biochemistry, 80, 557-85. [PubMed:21456964] [PMC] [WorldCat] [DOI] - ↑
Fitzpatrick, A.W., Debelouchina, G.T., Bayro, M.J., Clare, D.K., Caporini, M.A., Bajaj, V.S., ..., & Dobson, C.M. (2013).
Atomic structure and hierarchical assembly of a cross-β amyloid fibril. Proceedings of the National Academy of Sciences of the United States of America, 110(14), 5468-73. [PubMed:23513222] [PMC] [WorldCat] [DOI] - ↑
Klunk, W.E., Engler, H., Nordberg, A., Wang, Y., Blomqvist, G., Holt, D.P., ..., & Långström, B. (2004).
Imaging brain amyloid in Alzheimer's disease with Pittsburgh Compound-B. Annals of neurology, 55(3), 306-19. [PubMed:14991808] [WorldCat] [DOI] - ↑
Clark, C.M., Schneider, J.A., Bedell, B.J., Beach, T.G., Bilker, W.B., Mintun, M.A., ..., & AV45-A07 Study Group (2011).
Use of florbetapir-PET for imaging beta-amyloid pathology. JAMA, 305(3), 275-83. [PubMed:21245183] [WorldCat] [DOI] - ↑
Lee, Y.H., & Goto, Y. (2012).
Kinetic intermediates of amyloid fibrillation studied by hydrogen exchange methods with nuclear magnetic resonance. Biochimica et biophysica acta, 1824(12), 1307-23. [PubMed:22885025] [WorldCat] [DOI] - ↑
Jarrett, J.T., & Lansbury, P.T. (1993).
Seeding "one-dimensional crystallization" of amyloid: a pathogenic mechanism in Alzheimer's disease and scrapie? Cell, 73(6), 1055-8. [PubMed:8513491] [WorldCat] [DOI] - ↑
Nussbaum, J.M., Schilling, S., Cynis, H., Silva, A., Swanson, E., Wangsanut, T., ..., & Bloom, G.S. (2012).
Prion-like behaviour and tau-dependent cytotoxicity of pyroglutamylated amyloid-β. Nature, 485(7400), 651-5. [PubMed:22660329] [PMC] [WorldCat] [DOI] - ↑
Jucker, M., & Walker, L.C. (2013).
Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature, 501(7465), 45-51. [PubMed:24005412] [PMC] [WorldCat] [DOI] - ↑
Zhang, B., Une, Y., Fu, X., Yan, J., Ge, F., Yao, J., ..., & Higuchi, K. (2008).
Fecal transmission of AA amyloidosis in the cheetah contributes to high incidence of disease. Proceedings of the National Academy of Sciences of the United States of America, 105(20), 7263-8. [PubMed:18474855] [PMC] [WorldCat] [DOI] - ↑
Aguzzi, A., Nuvolone, M., & Zhu, C. (2013).
The immunobiology of prion diseases. Nature reviews. Immunology, 13(12), 888-902. [PubMed:24189576] [WorldCat] [DOI] - ↑
Eisele, Y.S., Obermüller, U., Heilbronner, G., Baumann, F., Kaeser, S.A., Wolburg, H., ..., & Jucker, M. (2010).
Peripherally applied Abeta-containing inoculates induce cerebral beta-amyloidosis. Science (New York, N.Y.), 330(6006), 980-2. [PubMed:20966215] [PMC] [WorldCat] [DOI] - ↑
Jucker, M., & Walker, L.C. (2013).
Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature, 501(7465), 45-51. [PubMed:24005412] [PMC] [WorldCat] [DOI] - ↑
Wickner, R.B., Edskes, H.K., Bateman, D.A., Kelly, A.C., Gorkovskiy, A., Dayani, Y., & Zhou, A. (2013).
Amyloids and yeast prion biology. Biochemistry, 52(9), 1514-27. [PubMed:23379365] [WorldCat] [DOI] - ↑
Lu, J.X., Qiang, W., Yau, W.M., Schwieters, C.D., Meredith, S.C., & Tycko, R. (2013).
Molecular structure of β-amyloid fibrils in Alzheimer's disease brain tissue. Cell, 154(6), 1257-68. [PubMed:24034249] [PMC] [WorldCat] [DOI] - ↑
Kayed, R., Head, E., Thompson, J.L., McIntire, T.M., Milton, S.C., Cotman, C.W., & Glabe, C.G. (2003).
Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science (New York, N.Y.), 300(5618), 486-9. [PubMed:12702875] [WorldCat] [DOI] - ↑
Cheng, B., Gong, H., Xiao, H., Petersen, R.B., Zheng, L., & Huang, K. (2013).
Inhibiting toxic aggregation of amyloidogenic proteins: a therapeutic strategy for protein misfolding diseases. Biochimica et biophysica acta, 1830(10), 4860-71. [PubMed:23820032] [WorldCat] [DOI] - ↑
Coomaraswamy, J., Kilger, E., Wölfing, H., Schäfer, C., Kaeser, S.A., Wegenast-Braun, B.M., ..., & Jucker, M. (2010).
Modeling familial Danish dementia in mice supports the concept of the amyloid hypothesis of Alzheimer's disease. Proceedings of the National Academy of Sciences of the United States of America, 107(17), 7969-74. [PubMed:20385796] [PMC] [WorldCat] [DOI]