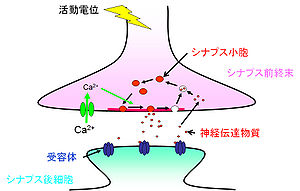
神経細胞はシナプスという構造を介して情報の伝達を行っている<ref>'''D Purves, GJ Augustine, D Fitzpatrick, WC Hall, AS LaMantia, JO McNamara, and LE White'''<br>Neuroscience, Fourth Edition<br>''Sinauer'':2011</ref>。神経細胞は情報を受容する細胞体と[[樹状突起]]、情報を出力する軸索から成る。軸索のシナプス結合部はやや膨大しており、シナプス前終末(presynaptic terminal)と呼ばれる。[[グルタミン酸]]を神経伝達物質とする[[興奮性]]シナプスでは、シナプス前終末からシナプス後細胞の樹状突起上のスパインへ情報が伝えられる(図1)。一方、GABAを伝達物質とする[[抑制性シナプス]]前終末は、樹状突起の幹および細胞体に接合する。シナプス前細胞と後細胞の間にはシナプス間隙(20nm)があり、情報伝達の場を形成している。その中には数百のシナプス小胞(50nm)が存在する。シナプス小胞がシナプス前終末の細胞膜と融合し、その中の神経伝達物質を開口放出([[エクソサイトーシス]];exocytosis)する領域は[[アクティブゾーン]]と呼ばれる。アクティブゾーンは5-20μm<sup>2</sup>の広さで、膜が肥厚しているように見え、5-10個のシナプス小胞が結合している。
神経細胞はシナプスという構造を介して情報の伝達を行っている<ref>'''D Purves, GJ Augustine, D Fitzpatrick, WC Hall, AS LaMantia, JO McNamara, and LE White'''<br>Neuroscience, Fourth Edition<br>''Sinauer'':2011<br></ref>。神経細胞は情報を受容する[[細胞体]]と[[樹状突起]]、情報を出力する軸索から成る。軸索のシナプス結合部はやや膨大しており、シナプス前終末と呼ばれる。[[グルタミン酸]]を神経伝達物質とする[[興奮性シナプス]]では、シナプス前終末からシナプス後細胞の樹状突起上の[[スパイン]]へ情報が伝えられる(図1)。一方、[[GABA]]を伝達物質とする[[抑制性シナプス]]前終末は、樹状突起の幹および細胞体に接合する。シナプス前細胞と後細胞の間にはシナプス間隙(20 nm)があり、情報伝達の場を形成している。
シナプス小胞は、その状態に応じて、[[即時放出可能プール]](readily releasable pool, RRP)・[[再循環プール]](recycling pool)・[[静止プール]](resting pool, RP)に分類される。即時放出可能プールと再循環プールを合わせて、[[全放出可能プール]]あるいは[[全リサイクリングプール]](total recycling pool, TRP)と呼ばれる。また、再循環プールと静止プールを[[貯蔵プール]](reserve pool)と呼ぶこともある。
N-ethylmaleimide-sensitive fusion protein (NSF)とsoluble NSF attachment protein (SNAP)は、ゴルジ体膜間の小胞輸送に必須な細胞質タンパク質である。これらのタンパク質は、2つの対峙した生体膜上に存在する受容体であるSNARE(SNAP receptor)タンパク質と相互作用することで、膜の融合を促すと考えられた。SNAREタンパク質は、融合する双方の膜表面に分かれた形で存在し、複合体を形成することによって膜間の距離を縮め、その結果として膜融合が引き起こされるというのがSNARE仮説である。一方を小胞、他方を細胞膜と想定し、それらの膜に存在するSNAREタンパク質は、各々v-SNARE、t-SNAREと名づけられた。シナプス前終末においては、シナプス小胞上のシナプトブレビン(VAMP; vesicle-associated membrane protein)がv-SNAREであり、アクティブゾーン細胞膜にあるシンタキシンとSNAP-25(synaptosomal associated protein-25)がt-SNAREに相当する<ref><pubmed> 11031229 </pubmed></ref>。[[SNARE複合体]]が、シナプスでの開口放出に中心的かつ必須な役割を果たすが、個々のSNAREタンパク質やSNARE複合体に相互作用するさまざまなタンパク質群が、いかに開口放出の時間的空間的制御に関わっているのかはまだよくわからない。前述したシナプス小胞上のシナプトタグミンは、エキソサイトーシスのカルシウムセンサーとしてはたらいて開口放出を制御するが、そのメカニズムは明確ではない<ref><pubmed> 21439657 </pubmed></ref>。<br>
シナプス後細胞から放出される[[逆行性伝達物質]]によっても、シナプス前終末からの伝達物質放出は制御される<ref><pubmed>19640475</pubmed></ref>。逆行性シナプス伝達制御の1つに[[Depolarization-induced suppression of inhibition|DSI]] ([[Depolarization-induced suppression of inhibition]])がある。DSIは、シナプス後細胞が脱分極することにより、そこから[[エンドカンナビノイド]]が放出され、抑制性シナプス前終末にある[[カンナビノイド受容体]]に作用することで、シナプス前終末からのシナプス小胞の放出確率が低下する現象である。
==関連項目==
*[[シナプス]]
*[[神経伝達物質]]
*[[シナプス小胞]]
*[[カルシウムチャネル]]
*[[トランスポーター]]
*[[活動電位]]
*[[エクソサイトーシス]]
*[[アクティブゾーン]]
*[[RIM1]]
*[[シナプトタグミン]]
*[[放出可能プール]]
*[[放出確率]]
*[[ニューレキシン]]
*[[シナプス前抑制]]
*[[Depolarization-induced suppression of inhibition]]
↑Iwasaki, S., Momiyama, A., Uchitel, O.D., & Takahashi, T. (2000).
Developmental changes in calcium channel types mediating central synaptic transmission. The Journal of neuroscience : the official journal of the Society for Neuroscience, 20(1), 59-65.
[PubMed:10627581]
[PMC]
[WorldCat]
↑Omote, H., Miyaji, T., Juge, N., & Moriyama, Y. (2011).
Vesicular neurotransmitter transporter: bioenergetics and regulation of glutamate transport. Biochemistry, 50(25), 5558-65.
[PubMed:21612282]
[WorldCat]
[DOI]
↑Südhof, T.C., & Rizo, J. (2011).
Synaptic vesicle exocytosis. Cold Spring Harbor perspectives in biology, 3(12).
[PubMed:22026965]
[PMC]
[WorldCat]
[DOI]
↑Lin, R.C., & Scheller, R.H. (2000).
Mechanisms of synaptic vesicle exocytosis. Annual review of cell and developmental biology, 16, 19-49.
[PubMed:11031229]
[WorldCat]
[DOI]
↑Kochubey, O., Lou, X., & Schneggenburger, R. (2011).
Regulation of transmitter release by Ca(2+) and synaptotagmin: insights from a large CNS synapse. Trends in neurosciences, 34(5), 237-46.
[PubMed:21439657]
[WorldCat]
[DOI]