平滑筋において、ノルアドレナリンα1受容体の活性化は、Gqを介してphospholipaseの活性化、inositol 1,4,5-trisphosphate (IP3)とdiacylglycerolの産生、IP3受容体の活性化による細胞内カルシウムストアからのカルシウム放出、myosin-light chain kinaseの活性化、そして結果的に筋収縮を引き起こす(<ref>'''D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong'''<br> Principles of Pharmacology, Second Edition<br>''Wolters Kluwer Health (Philadelphia)'':2002</ref>) (PMID: 11096123)。<br>逆に、アドレナリンによるβ2受容体の活性化は、Gsを介してPKAの活性化、MLCKのリン酸化による抑制の結果、筋弛緩をもたらすと考えられる(<ref>'''D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong'''<br> Principles of Pharmacology, Second Edition<br>''Wolters Kluwer Health (Philadelphia)'':2002</ref>)(PMID: 6259152)。<br>
こうした薬の作用から、うつ状態の原因がセロトニンやノルアドレナリンなどのモノアミンの減少によるのではないかという[[モノアミン仮説]]が生まれた。しかし、これらの薬の治療効果が現れるのは、モノアミン神経伝達が亢進されるよりもずっと遅いことから、この仮説よりももっと複雑なことが起きていると考えられている<ref name="ref20">'''E R Kandel, J H Schwartz, T M Jessell'''<br> Principles of Neural Science, Fourth Edition<br>''Mc Graw Hill (New York)'':2000</ref> <ref name="ref21">'''N R Carlson'''<br> Physiology of Behavior, Tenth Edition<br>''Pearson Education (Boston)'':2009</ref>。
こうした薬の作用から、うつ状態の原因がセロトニンやノルアドレナリンなどのモノアミンの減少によるのではないかという[[モノアミン仮説]]が生まれた。しかし、これらの薬の治療効果が現れるのは、モノアミン神経伝達が亢進されるよりもずっと遅いことから、この仮説よりももっと複雑なことが起きていると考えられている<ref name="ref58">'''E R Kandel, J H Schwartz, T M Jessell'''<br> Principles of Neural Science, Fourth Edition<br>''Mc Graw Hill (New York)'':2000</ref> <ref name="ref59">'''N R Carlson'''<br> Physiology of Behavior, Tenth Edition<br>''Pearson Education (Boston)'':2009</ref>。
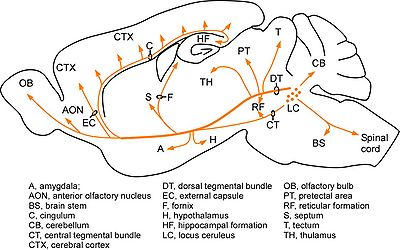
A6:青斑核(locus ceruleus)と呼ばれる。橋の背側に位置し、最も主要なノルアドレナリン作動性神経細胞の核である。青斑核からは、大脳皮質、視床、視床下部、小脳、中脳、脊髄、など脳のほぼ全域にわたって投射している(図2:Moore and Bloom, 1979[32]より許可を得て改変)。
↑U S von Euler A Specific Sympathomimetic Ergone in Adrenergic Nerve Fibres (Sympathin) and its Relations to Adrenaline and Nor-Adrenaline Acta Physiol., Scand. :1946, 12; 73–97
↑P Holtz Über die sympathicomimetische Wirksamkeit von Gehirnextrakten. Acta Physiol., Scand. : 1950, 20; 354–362
↑Dunkley, P.R., Bobrovskaya, L., Graham, M.E., von Nagy-Felsobuki, E.I., & Dickson, P.W. (2004).
Tyrosine hydroxylase phosphorylation: regulation and consequences. Journal of neurochemistry, 91(5), 1025-43.
[PubMed:15569247]
[WorldCat]
[DOI]
↑Daubner, S.C., Le, T., & Wang, S. (2011).
Tyrosine hydroxylase and regulation of dopamine synthesis. Archives of biochemistry and biophysics, 508(1), 1-12.
[PubMed:21176768]
[PMC]
[WorldCat]
[DOI]
↑Nagatsu, T. (1989).
The human tyrosine hydroxylase gene. Cellular and molecular neurobiology, 9(3), 313-21.
[PubMed:2575455]
[WorldCat]
[DOI]
↑Thöny, B., Auerbach, G., & Blau, N. (2000).
Tetrahydrobiopterin biosynthesis, regeneration and functions. The Biochemical journal, 347 Pt 1, 1-16.
[PubMed:10727395]
[PMC]
[WorldCat]
↑Rush, R.A., & Geffen, L.B. (1980).
Dopamine beta-hydroxylase in health and disease. Critical reviews in clinical laboratory sciences, 12(3), 241-77.
[PubMed:6998654]
[WorldCat]
[DOI]
↑Erickson, J.D., & Varoqui, H. (2000).
Molecular analysis of vesicular amine transporter function and targeting to secretory organelles. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 14(15), 2450-8.
[PubMed:11099462]
[WorldCat]
[DOI]
↑Mandela, P., & Ordway, G.A. (2006).
The norepinephrine transporter and its regulation. Journal of neurochemistry, 97(2), 310-33.
[PubMed:16539676]
[WorldCat]
[DOI]
↑ 14.014.1Youdim, M.B., Edmondson, D., & Tipton, K.F. (2006).
The therapeutic potential of monoamine oxidase inhibitors. Nature reviews. Neuroscience, 7(4), 295-309.
[PubMed:16552415]
[WorldCat]
[DOI]
↑Vitalis, T., Fouquet, C., Alvarez, C., Seif, I., Price, D., Gaspar, P., & Cases, O. (2002).
Developmental expression of monoamine oxidases A and B in the central and peripheral nervous systems of the mouse. The Journal of comparative neurology, 442(4), 331-47.
[PubMed:11793338]
[WorldCat]
[DOI]
↑Chen, J., Song, J., Yuan, P., Tian, Q., Ji, Y., Ren-Patterson, R., ..., & Weinberger, D.R. (2011).
Orientation and cellular distribution of membrane-bound catechol-O-methyltransferase in cortical neurons: implications for drug development. The Journal of biological chemistry, 286(40), 34752-60.
[PubMed:21846718]
[PMC]
[WorldCat]
[DOI]
↑ 17.017.117.217.3D E Golan, A H Tashjian Jr, E J Armstrong, A W Armstrong Principles of Pharmacology, Second Edition Wolters Kluwer Health (Philadelphia):2002
↑Madison, D.V., & Nicoll, R.A. (1986).
Actions of noradrenaline recorded intracellularly in rat hippocampal CA1 pyramidal neurones, in vitro. The Journal of physiology, 372, 221-44.
[PubMed:2873241]
[PMC]
[WorldCat]
[DOI]
↑Haas, H.L., & Konnerth, A. (1983).
Histamine and noradrenaline decrease calcium-activated potassium conductance in hippocampal pyramidal cells. Nature, 302(5907), 432-4.
[PubMed:6300681]
[WorldCat]
[DOI]
↑Pedarzani, P., & Storm, J.F. (1993).
PKA mediates the effects of monoamine transmitters on the K+ current underlying the slow spike frequency adaptation in hippocampal neurons. Neuron, 11(6), 1023-35.
[PubMed:8274274]
[WorldCat]
[DOI]
↑Neuman, R.S., & Harley, C.W. (1983).
Long-lasting potentiation of the dentate gyrus population spike by norepinephrine. Brain research, 273(1), 162-5.
[PubMed:6311345]
[WorldCat]
[DOI]
↑O'Dell, T.J., Connor, S.A., Gelinas, J.N., & Nguyen, P.V. (2010).
Viagra for your synapses: Enhancement of hippocampal long-term potentiation by activation of beta-adrenergic receptors. Cellular signalling, 22(5), 728-36.
[PubMed:20043991]
[PMC]
[WorldCat]
[DOI]
↑Hoffman, D.A., & Johnston, D. (1999).
Neuromodulation of dendritic action potentials. Journal of neurophysiology, 81(1), 408-11.
[PubMed:9914302]
[WorldCat]
[DOI]
↑Yuan, L.L., Adams, J.P., Swank, M., Sweatt, J.D., & Johnston, D. (2002).
Protein kinase modulation of dendritic K+ channels in hippocampus involves a mitogen-activated protein kinase pathway. The Journal of neuroscience : the official journal of the Society for Neuroscience, 22(12), 4860-8.
[PubMed:12077183]
[PMC]
[WorldCat]
↑Sampson, K.J., & Kass, R.S. (2010).
Molecular mechanisms of adrenergic stimulation in the heart. Heart rhythm, 7(8), 1151-3.
[PubMed:20156590]
[PMC]
[WorldCat]
[DOI]
↑Wahl-Schott, C., & Biel, M. (2009).
HCN channels: structure, cellular regulation and physiological function. Cellular and molecular life sciences : CMLS, 66(3), 470-94.
[PubMed:18953682]
[WorldCat]
[DOI]
↑ 27.027.1Sampson, K.J., & Kass, R.S. (2010).
Molecular mechanisms of adrenergic stimulation in the heart. Heart rhythm, 7(8), 1151-3.
[PubMed:20156590]
[PMC]
[WorldCat]
[DOI]
↑Gray, P.C., Scott, J.D., & Catterall, W.A. (1998).
Regulation of ion channels by cAMP-dependent protein kinase and A-kinase anchoring proteins. Current opinion in neurobiology, 8(3), 330-4.
[PubMed:9687361]
[WorldCat]
[DOI]
↑Kamm, K.E., & Stull, J.T. (2001).
Dedicated myosin light chain kinases with diverse cellular functions. The Journal of biological chemistry, 276(7), 4527-30.
[PubMed:11096123]
[WorldCat]
[DOI]
↑Conti, M.A., & Adelstein, R.S. (1981).
The relationship between calmodulin binding and phosphorylation of smooth muscle myosin kinase by the catalytic subunit of 3':5' cAMP-dependent protein kinase. The Journal of biological chemistry, 256(7), 3178-81.
[PubMed:6259152]
[WorldCat]
↑Starke, K. (2001).
Presynaptic autoreceptors in the third decade: focus on alpha2-adrenoceptors. Journal of neurochemistry, 78(4), 685-93.
[PubMed:11520889]
[WorldCat]
[DOI]
↑Moore, R.Y., & Bloom, F.E. (1978).
Central catecholamine neuron systems: anatomy and physiology of the dopamine systems. Annual review of neuroscience, 1, 129-69.
[PubMed:756202]
[WorldCat]
[DOI]
↑Moruzzi, G., & Magoun, H.W. (1949).
Brain stem reticular formation and activation of the EEG. Electroencephalography and clinical neurophysiology, 1(4), 455-73.
[PubMed:18421835]
[WorldCat]
↑Aston-Jones, G., & Bloom, F.E. (1981).
Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. The Journal of neuroscience : the official journal of the Society for Neuroscience, 1(8), 876-86.
[PubMed:7346592]
[WorldCat]
↑Foote, S.L., Aston-Jones, G., & Bloom, F.E. (1980).
Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proceedings of the National Academy of Sciences of the United States of America, 77(5), 3033-7.
[PubMed:6771765]
[PMC]
[WorldCat]
[DOI]
↑Foote, S.L., Freedman, R., & Oliver, A.P. (1975).
Effects of putative neurotransmitters on neuronal activity in monkey auditory cortex. Brain research, 86(2), 229-42.
[PubMed:234774]
[WorldCat]
[DOI]
↑Waterhouse, B.D., & Woodward, D.J. (1980).
Interaction of norepinephrine with cerebrocortical activity evoked by stimulation of somatosensory afferent pathways in the rat. Experimental neurology, 67(1), 11-34.
[PubMed:7349980]
[WorldCat]
[DOI]
↑Segal, M., & Bloom, F.E. (1976).
The action of norepinephrine in the rat hippocampus. IV. The effects of locus coeruleus stimulation on evoked hippocampal unit activity. Brain research, 107(3), 513-25.
[PubMed:178411]
[WorldCat]
[DOI]
↑Lecas, J.C. (2004).
Locus coeruleus activation shortens synaptic drive while decreasing spike latency and jitter in sensorimotor cortex. Implications for neuronal integration. The European journal of neuroscience, 19(9), 2519-30.
[PubMed:15128405]
[WorldCat]
[DOI]
↑Bouret, S., & Sara, S.J. (2002).
Locus coeruleus activation modulates firing rate and temporal organization of odour-induced single-cell responses in rat piriform cortex. The European journal of neuroscience, 16(12), 2371-82.
[PubMed:12492432]
[WorldCat]
[DOI]
↑Mair, R.D., Zhang, Y., Bailey, K.R., Toupin, M.M., & Mair, R.G. (2005).
Effects of clonidine in the locus coeruleus on prefrontal- and hippocampal-dependent measures of attention and memory in the rat. Psychopharmacology, 181(2), 280-8.
[PubMed:15830223]
[WorldCat]
[DOI]
↑Ramos, B.P., & Arnsten, A.F. (2007).
Adrenergic pharmacology and cognition: focus on the prefrontal cortex. Pharmacology & therapeutics, 113(3), 523-36.
[PubMed:17303246]
[PMC]
[WorldCat]
[DOI]
↑Wang, M., Ramos, B.P., Paspalas, C.D., Shu, Y., Simen, A., Duque, A., ..., & Arnsten, A.F. (2007).
Alpha2A-adrenoceptors strengthen working memory networks by inhibiting cAMP-HCN channel signaling in prefrontal cortex. Cell, 129(2), 397-410.
[PubMed:17448997]
[WorldCat]
[DOI]
↑Coull, J.T., Büchel, C., Friston, K.J., & Frith, C.D. (1999).
Noradrenergically mediated plasticity in a human attentional neuronal network. NeuroImage, 10(6), 705-15.
[PubMed:10600416]
[WorldCat]
[DOI]
↑McGaugh, J.L., & Roozendaal, B. (2009).
Drug enhancement of memory consolidation: historical perspective and neurobiological implications. Psychopharmacology, 202(1-3), 3-14.
[PubMed:18704369]
[WorldCat]
[DOI]
↑Sara, S.J., Roullet, P., & Przybyslawski, J. (1999).
Consolidation of memory for odor-reward association: beta-adrenergic receptor involvement in the late phase. Learning & memory (Cold Spring Harbor, N.Y.), 6(2), 88-96.
[PubMed:10327234]
[PMC]
[WorldCat]
↑Tronel, S., Feenstra, M.G., & Sara, S.J. (2004).
Noradrenergic action in prefrontal cortex in the late stage of memory consolidation. Learning & memory (Cold Spring Harbor, N.Y.), 11(4), 453-8.
[PubMed:15254217]
[PMC]
[WorldCat]
[DOI]
↑Sara, S.J., & Devauges, V. (1988).
Priming stimulation of locus coeruleus facilitates memory retrieval in the rat. Brain research, 438(1-2), 299-303.
[PubMed:3345434]
[WorldCat]
[DOI]
↑Sara, S.J., & Devauges, V. (1989).
Idazoxan, an alpha-2 antagonist, facilitates memory retrieval in the rat. Behavioral and neural biology, 51(3), 401-11.
[PubMed:2543356]
[WorldCat]
[DOI]
↑Murchison, C.F., Zhang, X.Y., Zhang, W.P., Ouyang, M., Lee, A., & Thomas, S.A. (2004).
A distinct role for norepinephrine in memory retrieval. Cell, 117(1), 131-43.
[PubMed:15066288]
[WorldCat]
[DOI]
↑ 52.052.1Strawn, J.R., & Geracioti, T.D. (2008).
Noradrenergic dysfunction and the psychopharmacology of posttraumatic stress disorder. Depression and anxiety, 25(3), 260-71.
[PubMed:17354267]
[WorldCat]
[DOI]
↑Debiec, J., & LeDoux, J.E. (2006).
Noradrenergic signaling in the amygdala contributes to the reconsolidation of fear memory: treatment implications for PTSD. Annals of the New York Academy of Sciences, 1071, 521-4.
[PubMed:16891611]
[WorldCat]
[DOI]
↑Diergaarde, L., Schoffelmeer, A.N., & De Vries, T.J. (2008).
Pharmacological manipulation of memory reconsolidation: towards a novel treatment of pathogenic memories. European journal of pharmacology, 585(2-3), 453-7.
[PubMed:18410917]
[WorldCat]
[DOI]
↑Brunet, A., Orr, S.P., Tremblay, J., Robertson, K., Nader, K., & Pitman, R.K. (2008).
Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumatic stress disorder. Journal of psychiatric research, 42(6), 503-6.
[PubMed:17588604]
[WorldCat]
[DOI]
↑E R Kandel, J H Schwartz, T M Jessell Principles of Neural Science, Fourth Edition Mc Graw Hill (New York):2000
↑N R Carlson Physiology of Behavior, Tenth Edition Pearson Education (Boston):2009