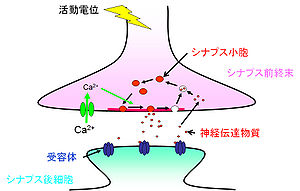
神経細胞はシナプスという構造を介して情報の伝達を行っている<ref>'''D Purves, GJ Augustine, D Fitzpatrick, WC Hall, AS LaMantia, JO McNamara, and LE White'''<br><br>Neuroscience Fourth Edition<br><br>''Sinauer'':2011</ref>。神経細胞は情報を受容する細胞体と[[樹状突起]]、情報を出力する軸索から成る。軸索のシナプス結合部はやや膨大しており、シナプス前終末(presynaptic terminal)と呼ばれる。[[グルタミン酸]]を神経伝達物質とする[[興奮性]]シナプスでは、シナプス前終末からシナプス後細胞の樹状突起上のスパインへ情報が伝えられる(図1)。一方、GABAを伝達物質とする[[抑制性シナプス]]前終末は、樹状突起の幹および細胞体に接合する。シナプス前細胞と後細胞の間にはシナプス間隙(20nm)があり、情報伝達の場を形成している。その中には数百のシナプス小胞(50nm)が存在する。シナプス小胞がシナプス前終末の細胞膜と融合し、その中の神経伝達物質を開口放出([[エクソサイトーシス]];exocytosis)する領域は[[アクティブゾーン]]と呼ばれる。アクティブゾーンは5-20μm<sup>2</sup>の広さで、膜が肥厚しているように見え、5-10個のシナプス小胞が結合している。
神経細胞はシナプスという構造を介して情報の伝達を行っている<ref>'''D Purves, GJ Augustine, D Fitzpatrick, WC Hall, AS LaMantia, JO McNamara, and LE White'''<br>Neuroscience, Fourth Edition<br>''Sinauer'':2011</ref>。神経細胞は情報を受容する細胞体と[[樹状突起]]、情報を出力する軸索から成る。軸索のシナプス結合部はやや膨大しており、シナプス前終末(presynaptic terminal)と呼ばれる。[[グルタミン酸]]を神経伝達物質とする[[興奮性]]シナプスでは、シナプス前終末からシナプス後細胞の樹状突起上のスパインへ情報が伝えられる(図1)。一方、GABAを伝達物質とする[[抑制性シナプス]]前終末は、樹状突起の幹および細胞体に接合する。シナプス前細胞と後細胞の間にはシナプス間隙(20nm)があり、情報伝達の場を形成している。その中には数百のシナプス小胞(50nm)が存在する。シナプス小胞がシナプス前終末の細胞膜と融合し、その中の神経伝達物質を開口放出([[エクソサイトーシス]];exocytosis)する領域は[[アクティブゾーン]]と呼ばれる。アクティブゾーンは5-20μm<sup>2</sup>の広さで、膜が肥厚しているように見え、5-10個のシナプス小胞が結合している。
↑Iwasaki, S., Momiyama, A., Uchitel, O.D., & Takahashi, T. (2000).
Developmental changes in calcium channel types mediating central synaptic transmission. The Journal of neuroscience : the official journal of the Society for Neuroscience, 20(1), 59-65.
[PubMed:10627581]
[PMC]
[WorldCat]
↑Omote, H., Miyaji, T., Juge, N., & Moriyama, Y. (2011).
Vesicular neurotransmitter transporter: bioenergetics and regulation of glutamate transport. Biochemistry, 50(25), 5558-65.
[PubMed:21612282]
[WorldCat]
[DOI]
↑Südhof, T.C., & Rizo, J. (2011).
Synaptic vesicle exocytosis. Cold Spring Harbor perspectives in biology, 3(12).
[PubMed:22026965]
[PMC]
[WorldCat]
[DOI]
↑Lin, R.C., & Scheller, R.H. (2000).
Mechanisms of synaptic vesicle exocytosis. Annual review of cell and developmental biology, 16, 19-49.
[PubMed:11031229]
[WorldCat]
[DOI]
↑Kochubey, O., Lou, X., & Schneggenburger, R. (2011).
Regulation of transmitter release by Ca(2+) and synaptotagmin: insights from a large CNS synapse. Trends in neurosciences, 34(5), 237-46.
[PubMed:21439657]
[WorldCat]
[DOI]