「P1受容体」の版間の差分
細編集の要約なし |
細編集の要約なし |
||
| (同じ利用者による、間の4版が非表示) | |||
| 2行目: | 2行目: | ||
<font size="+1">[http://yakkou.phar.kyushu-u.ac.jp/Member.html 津田 誠]</font><br> | <font size="+1">[http://yakkou.phar.kyushu-u.ac.jp/Member.html 津田 誠]</font><br> | ||
''九州大学大学院薬学研究院 医療薬科学部門 薬理学分野''<br> | ''九州大学大学院薬学研究院 医療薬科学部門 薬理学分野''<br> | ||
DOI:<selfdoi /> 原稿受付日:2013年6月14日 原稿完成日:2015年12月7日<br> | |||
担当編集委員:[http://researchmap.jp/2rikenbsi 林 康紀](独立行政法人理化学研究所 脳科学総合研究センター)<br> | 担当編集委員:[http://researchmap.jp/2rikenbsi 林 康紀](独立行政法人理化学研究所 脳科学総合研究センター)<br> | ||
</div> | </div> | ||
| 10行目: | 10行目: | ||
同義語:アデノシン受容体 | 同義語:アデノシン受容体 | ||
{{box|text= | {{box|text= P1受容体とは、アデノシンをリガンドとする一群の細胞表面受容体である。Gタンパク質共役型受容体(GPCR)で、A<sub>1</sub>、A<sub>2A</sub>、A<sub>2B</sub>およびA<sub>3</sub>に分類される。}} | ||
{{PBB|geneid=134}} | {{PBB|geneid=134}} | ||
{{PBB|geneid=135}} | {{PBB|geneid=135}} | ||
| 16行目: | 16行目: | ||
{{PBB|geneid=140}} | {{PBB|geneid=140}} | ||
== P1受容体とは== | == P1受容体とは== | ||
P1受容体とは、[[アデノシン]]を[[リガンド]]とする一群の細胞表面[[受容体]]である。[[Gタンパク質共役型受容体]](GPCR)で、[[A1受容体| | P1受容体とは、[[アデノシン]]を[[リガンド]]とする一群の細胞表面[[受容体]]である。[[Gタンパク質共役型受容体]](GPCR)で、[[A1受容体|A<sub>1</sub>]]、[[A2A受容体|A<sub>2A</sub>]]、[[A2B受容体|A<sub>2B</sub>]]および[[A3受容体|A<sub>3</sub>]]に分類される。[[プリンヌクレオチド]]([[ATP]]、[[ADP]])、[[ピリミジンヌクレオチド]]([[UTP]]、[[UDP]])、[[糖ヌクレオチド]]などを内因性リガンドとする[[P2受容体]]と共に[[プリン受容体]]と呼ばれる('''表1、2''')<ref name=ref1><pubmed>17429044</pubmed></ref> <ref name=ref2><pubmed>18591979</pubmed></ref>。 | ||
なお、本項における受容体の表記は、IUPHAR [http://www.iuphar-db.org/index.jsp 国際薬理学連合]でのデータベース掲載名に従った。 | なお、本項における受容体の表記は、IUPHAR [http://www.iuphar-db.org/index.jsp 国際薬理学連合]でのデータベース掲載名に従った。 | ||
| 24行目: | 24行目: | ||
|colspan="2"|'''名称''' ||'''下位分類'''|| '''リガンド''' || '''作用機構''' | |colspan="2"|'''名称''' ||'''下位分類'''|| '''リガンド''' || '''作用機構''' | ||
|- | |- | ||
| colspan="2"|[[P1受容体]] || | | colspan="2"|[[P1受容体]] ||A<sub>1</sub>、A<sub>2A</sub>、A<sub>2B</sub>,A<sub>3</sub>受容体|| [[アデノシン]] || [[Gタンパク質共役型受容体]] | ||
|- | |- | ||
| rowspan="2"|'''P2受容体'''||'''[[P2X受容体]]''' ||P2X1−7受容体||ATP || [[リガンド依存性イオンチャンネル]] | | rowspan="2"|'''P2受容体'''||'''[[P2X受容体]]''' ||P2X1−7受容体||ATP || [[リガンド依存性イオンチャンネル]] | ||
|- | |- | ||
|| '''[[P2Y受容体]]''' || | || '''[[P2Y受容体]]''' ||P2Y<sub>1, 2, 4, 6, 11−14</sub>受容体||[[ヌクレオチド]] | ||
* [[wj:アデノシン三リン酸|ATP]] | * [[wj:アデノシン三リン酸|ATP]] | ||
* [[wj:アデノシン二リン酸|ADP]] | * [[wj:アデノシン二リン酸|ADP]] | ||
| 37行目: | 37行目: | ||
|} | |} | ||
編集部にて[[w:Purinergic receptor|Wikipedia]]より翻訳、改変。 | 編集部にて[[w:Purinergic receptor|Wikipedia]]より翻訳、改変。 | ||
== サブタイプ == | == サブタイプ == | ||
=== | === A<sub>1</sub>受容体 === | ||
[[Giタンパク質|Gi]]/[[Goタンパク質]]共役型受容体である。[[カフェイン]]の作用標的としても知られている。[[脳]]や[[脊髄]]などの[[中枢神経系]]組織に加え、[[心臓]]や[[腎臓]]、[[肺]]といった末梢組織を含む全身の様々な組織に発現している<ref name=ref3><pubmed>8234299</pubmed></ref> <ref name=ref4><pubmed>18984166</pubmed></ref> | [[Giタンパク質|Gi]]/[[Goタンパク質]]共役型受容体である。[[カフェイン]]の作用標的としても知られている。[[脳]]や[[脊髄]]などの[[中枢神経系]]組織に加え、[[心臓]]や[[腎臓]]、[[肺]]といった末梢組織を含む全身の様々な組織に発現している<ref name=ref3><pubmed>8234299</pubmed></ref> <ref name=ref4><pubmed>18984166</pubmed></ref>。[[A1受容体|A<sub>1</sub>受容体]]は[[P2Y1受容体]]とヘテロマーを形成することも知られている<ref name=ref5><pubmed>12417330</pubmed></ref>。これまでに数十種類の選択的[[作動薬]]や[[拮抗薬]]が合成され、その機能解明も進んでいる。例えば、脳において[[興奮性シナプス]]伝達を抑制的に制御していることが知られており<ref name=ref6><pubmed>23332692</pubmed></ref>、A1受容体欠損マウスの[[海馬]]においては、シナプス伝達の異常興奮が観察される<ref name=ref7><pubmed>11470917</pubmed></ref>。また、[[心筋]]の収縮力や[[心拍数]]の制御に加え、[[血管]]拡張作用、[[体温調節]]機構にも関与していると考えられている<ref name=ref7 /> <ref name=ref8><pubmed>11641103</pubmed></ref>。A1受容体欠損マウスは、[[てんかん]]症状や[[痛覚]]過敏症状、[[不安]]関連行動を呈するほか、[[インスリン]][[分泌]]の亢進や低酸素性障害や[[虚血]][[ストレス]]に対する抵抗性の低下が観察される<ref name=ref7 /> <ref name=ref9><pubmed>15661449</pubmed></ref>。 | ||
=== | ===A<sub>2A</sub>受容体=== | ||
[[Gsタンパク質]]共役型GPCRで、受容体刺激により[[アデニル酸シクラーゼ]]が活性化され、[[cAMP]] | [[Gsタンパク質]]共役型GPCRで、受容体刺激により[[アデニル酸シクラーゼ]]が活性化され、[[cAMP]]の産生が亢進する。A<sub>1</sub>受容体<ref name=ref10><pubmed>16481441</pubmed></ref>や他の[[神経伝達物質]]受容体([[ドーパミン]][[D2受容体]]<ref name=ref11><pubmed>16012194</pubmed></ref>, [[D3受容体]]<ref name=ref12><pubmed>15539641</pubmed></ref>, [[代謝型グルタミン酸受容体]][[mGluR5]]<ref name=ref13><pubmed>18246094 </pubmed></ref>, [[CB1受容体]]<ref name=ref14><pubmed>18691604</pubmed></ref>)とヘテロマーを形成することがある。A<sub>1</sub>受容体と同様に、カフェインが拮抗薬として作用する。また、[[A2A受容体|A<sub>2A</sub>受容体]]に対する選択的作用薬として[[CGS21680]]や[[DPMA]]、拮抗薬として[[イストラデフィリン]]や[[SCH-58261]]がある<ref name=ref15><pubmed>22371149</pubmed></ref>。A<sub>2A</sub>受容体は、生体内に幅広く分布し、[[線条体]]や海馬、[[冠血管]]、肺、[[血小板]]、腎臓などに発現する<ref name=ref16><pubmed>9822147</pubmed></ref> <ref name=ref17><pubmed>8522976</pubmed></ref> <ref name=ref18><pubmed>8732278</pubmed></ref>。生理的役割としては、血管拡張、[[睡眠]]、神経活動制御があり、A<sub>2A</sub>受容体欠損マウスにおいて不安行動及び[[攻撃性]]増加や[[痛覚]]鈍麻、心拍数増加、血圧上昇、血小板凝集が見られる<ref name=ref19><pubmed>8650205</pubmed></ref> <ref name=ref20><pubmed>9262401</pubmed></ref>。最近、[[ミクログリア]]細胞にA<sub>2A</sub>受容体が発現し、突起の退縮に関与していることも報告された<ref name=ref21><pubmed>19525944</pubmed></ref>。現在、A<sub>2A</sub>受容体作動薬[[レガデノソン]](レキスキャン®、アステラス製薬/CVセラピューティクス)が心筋血流イメージングの薬物負荷剤として、A2A受容体拮抗薬イストラデフィリン(ノウリアスト®、協和発酵キリン)が[[パーキンソン病]]治療薬として用いられている。 | ||
=== | ===A<sub>2B</sub>受容体=== | ||
A<sub>2A</sub>受容体と同様にGs共役型GPCRであるが、アデノシンへの親和性が低く、アデノシンの他に[[ネトリン-1]]の受容体としても機能する<ref name=ref22><pubmed>11048721</pubmed></ref>。[[A2B受容体|A<sub>2B</sub>受容体]]選択的作動薬として[[BAY 60-6583]]や[[NECA]]、拮抗薬として[[MRS-1754]]や[[MRE-2029-F20]]、[[CVT6883]]、[[PSB-1115]]などがある<ref name=ref15 />。A<sub>2B</sub>受容体は、中枢神経系を含め、全身に広く発現しており<ref name=ref23><pubmed>9443164</pubmed></ref>、特に脳や腸、腎臓、肺に多く、心臓や[[大動脈]]にもわずかに発現が見られるが、[[肝臓]]ではほとんど見られない<ref name=ref24><pubmed>16823489</pubmed></ref>。また、[[肥満細胞]]や[[線維芽細胞]]にも発現する<ref name=ref23 /> <ref name=ref25><pubmed>17200408</pubmed></ref>。A2B受容体欠損マウスにおいて、[[リポ多糖]](LPS)による炎症及び[[サイトカイン]]産生の抑制や[[リンパ球]]の血管付着の増加<ref name=ref24 />、肥満細胞の活性化及び[[IgE]]誘発[[アナフィラキシー]]亢進<ref name=ref25 />、心筋虚血プレコンデショニングによる心臓保護作用の抑制が報告されている<ref name=ref26><pubmed>17353435</pubmed></ref>。 | |||
=== | ===A<sub>3</sub>受容体=== | ||
薬理学的機能同定前にクローニングされた唯一のアデノシン受容体サブタイプで、Gi、Goあるいは[[Gqタンパク質]]と共役し細胞内にシグナルを伝える<ref name=ref6 />。肺や腎臓、心臓、脳、脾臓、肝臓など全身の様々な組織に発現しているが、その発現レベルは動物種間で大きく異なっている<ref name=ref4 /> <ref name=ref27><pubmed>18029023</pubmed></ref>。例えば、[[ラット]]では[[睾丸]]や肥満細胞で発現量が高いのに対して、ヒトでは肺や肝臓でその発現が高く、脳や大動脈での発現量は低い<ref name=ref3 /> | 薬理学的機能同定前にクローニングされた唯一のアデノシン受容体サブタイプで、Gi、Goあるいは[[Gqタンパク質]]と共役し細胞内にシグナルを伝える<ref name=ref6 />。肺や腎臓、心臓、脳、脾臓、肝臓など全身の様々な組織に発現しているが、その発現レベルは動物種間で大きく異なっている<ref name=ref4 /> <ref name=ref27><pubmed>18029023</pubmed></ref>。例えば、[[ラット]]では[[睾丸]]や肥満細胞で発現量が高いのに対して、ヒトでは肺や肝臓でその発現が高く、脳や大動脈での発現量は低い<ref name=ref3 />。[[A3受容体|A<sub>3</sub>受容体]]欠損マウスにおいては、野生型マウスと比較して、眼圧の低下<ref name=ref28><pubmed>12202525</pubmed></ref>や局所炎症反応の減少<ref name=ref29><pubmed>12220556</pubmed></ref>など、いくつかの表現型の違いが観察される。 | ||
{| class="wikitable" | {| class="wikitable" | ||
|+表2. P1受容体(アデノシン受容体) | |+表2. P1受容体(アデノシン受容体) | ||
| 54行目: | 55行目: | ||
! 受容体 !! 遺伝子 !!Allen Brain Atlas!! 細胞内情報伝達機構 !! 作用 !! アゴニスト !! アンタゴニスト | ! 受容体 !! 遺伝子 !!Allen Brain Atlas!! 細胞内情報伝達機構 !! 作用 !! アゴニスト !! アンタゴニスト | ||
|- | |- | ||
| [[A1受容体|A<sub>1</sub>受容体]]|| {{Gene|ADORA1}} ||[http://mouse.brain-map.org/experiment/show/73496905 73496905]|| [[Giタンパク質|Gi]]/[[Goタンパク質|o]] → [[サイクリックAMP|cAMP]]↑/↓ | |||
| {{Gene|ADORA1}} ||[http://mouse.brain-map.org/experiment/show/73496905 73496905]|| [[Giタンパク質|Gi]]/[[Goタンパク質|o]] → [[サイクリックAMP|cAMP]]↑/↓ | |||
|| | || | ||
* 心拍数現象 | * 心拍数現象 | ||
| 77行目: | 77行目: | ||
*[[PSB 36]] | *[[PSB 36]] | ||
|- | |- | ||
| [[A2A受容体|A<sub>2A</sub>受容体]]|| {{Gene|ADORA2A}} ||[http://mouse.brain-map.org/experiment/show/69855739 69855739]|| [[Gsタンパク質|Gs]] → cAMP↑ || | |||
| {{Gene|ADORA2A}} ||[http://mouse.brain-map.org/experiment/show/69855739 69855739]|| [[Gsタンパク質|Gs]] → cAMP↑ || | |||
* [[冠動脈]]拡張 | * [[冠動脈]]拡張 | ||
* ドーパミン神経活動減弱 | * ドーパミン神経活動減弱 | ||
| 96行目: | 95行目: | ||
*[[ZM-241,385]] | *[[ZM-241,385]] | ||
|- | |- | ||
| [[A2B受容体|A<sub>2B</sub>受容体]]|| {{Gene|ADORA2B}} ||[http://mouse.brain-map.org/experiment/show/69855773 69855773]|| Gs → cAMP↑ | |||
| {{Gene|ADORA2B}} ||[http://mouse.brain-map.org/experiment/show/69855773 69855773]|| Gs → cAMP↑ | |||
|| | || | ||
*気管収縮 | *気管収縮 | ||
| 117行目: | 115行目: | ||
*[[PSB-1115]] | *[[PSB-1115]] | ||
|- | |- | ||
| [[A3受容体|A<sub>3</sub>受容体]]|| {{Gene|ADORA3}} ||[http://mouse.brain-map.org/experiment/show/75828188 75828188]|| [[Giタンパク質|Gi]] → cAMP↓|| | |||
| {{Gene|ADORA3}} ||[http://mouse.brain-map.org/experiment/show/75828188 75828188]|| [[Giタンパク質|Gi]] → cAMP↓|| | |||
*心筋弛緩 | *心筋弛緩 | ||
*平滑筋収縮 | *平滑筋収縮 | ||
2021年6月3日 (木) 18:06時点における版
津田 誠
九州大学大学院薬学研究院 医療薬科学部門 薬理学分野
DOI:10.14931/bsd.6532 原稿受付日:2013年6月14日 原稿完成日:2015年12月7日
担当編集委員:林 康紀(独立行政法人理化学研究所 脳科学総合研究センター)
英語名:P1 purinergic receptor 独:P1 purinerge Rezeptoren 仏:récepteur purinergique P1
同義語:アデノシン受容体
P1受容体とは、アデノシンをリガンドとする一群の細胞表面受容体である。Gタンパク質共役型受容体(GPCR)で、A1、A2A、A2BおよびA3に分類される。
{kind=link}
{kind=link}
| アデノシンA2A受容体 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
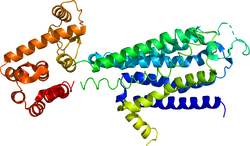 アデノシンA2A受容体の結晶構造。3EMLによる。 | |||||||||||||
| |||||||||||||
| Identifiers | |||||||||||||
| Symbols | ADORA2A; A2aR; ADORA2; RDC8 | ||||||||||||
| External IDs | OMIM: 102776 MGI: 99402 HomoloGene: 20166 IUPHAR: A2A GeneCards: ADORA2A Gene | ||||||||||||
| |||||||||||||
| RNA expression pattern | |||||||||||||
| ファイル:PBB GE ADORA2A 205013 s at tn.png | |||||||||||||
| More reference expression data | |||||||||||||
| Orthologs | |||||||||||||
| Species | Human | Mouse | |||||||||||
| Entrez | 135 | 11540 | |||||||||||
| Ensembl | ENSG00000128271 | ENSMUSG00000020178 | |||||||||||
| UniProt | P29274 | Q60613 | |||||||||||
| RefSeq (mRNA) | NM_000675 | NM_009630 | |||||||||||
| RefSeq (protein) | NP_000666 | NP_033760 | |||||||||||
| Location (UCSC) |
Chr 22: 24.42 – 24.44 Mb |
Chr 10: 75.32 – 75.33 Mb | |||||||||||
| PubMed search | [3] | [4] | |||||||||||
{kind=link}
| Adenosine A2b receptor | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||
| Symbols | ADORA2B; ADORA2 | ||||||||||||
| External IDs | OMIM: 600446 MGI: 99403 HomoloGene: 20167 IUPHAR: A2B GeneCards: ADORA2B Gene | ||||||||||||
| |||||||||||||
| RNA expression pattern | |||||||||||||
| ファイル:PBB GE ADORA2B 205891 at tn.png | |||||||||||||
| More reference expression data | |||||||||||||
| Orthologs | |||||||||||||
| Species | Human | Mouse | |||||||||||
| Entrez | 136 | 11541 | |||||||||||
| Ensembl | ENSG00000170425 | ENSMUSG00000018500 | |||||||||||
| UniProt | P29275 | Q60614 | |||||||||||
| RefSeq (mRNA) | NM_000676 | NM_007413 | |||||||||||
| RefSeq (protein) | NP_000667 | NP_031439 | |||||||||||
| Location (UCSC) |
Chr 17: 15.94 – 15.98 Mb |
Chr 11: 62.25 – 62.27 Mb | |||||||||||
| PubMed search | [5] | [6] | |||||||||||
{kind=link}
| Adenosine A3 receptor | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||
| Identifiers | |||||||||||||
| Symbols | ADORA3; A3AR | ||||||||||||
| External IDs | OMIM: 600445 MGI: 104847 HomoloGene: 550 IUPHAR: A3 GeneCards: ADORA3 Gene | ||||||||||||
| |||||||||||||
| RNA expression pattern | |||||||||||||
| ファイル:PBB GE ADORA3 206171 at tn.png | |||||||||||||
| More reference expression data | |||||||||||||
| Orthologs | |||||||||||||
| Species | Human | Mouse | |||||||||||
| Entrez | 140 | 11542 | |||||||||||
| Ensembl | ENSG00000282608 | ENSMUSG00000000562 | |||||||||||
| UniProt | P0DMS8 | Q3U4C5 | |||||||||||
| RefSeq (mRNA) | NM_000677 | NM_009631 | |||||||||||
| RefSeq (protein) | NP_000668 | NP_033761 | |||||||||||
| Location (UCSC) |
Chr 1: 111.5 – 111.5 Mb |
Chr 3: 105.87 – 105.91 Mb | |||||||||||
| PubMed search | [7] | [8] | |||||||||||
{kind=link}
P1受容体とは
P1受容体とは、アデノシンをリガンドとする一群の細胞表面受容体である。Gタンパク質共役型受容体(GPCR)で、A1、A2A、A2BおよびA3に分類される。プリンヌクレオチド(ATP、ADP)、ピリミジンヌクレオチド(UTP、UDP)、糖ヌクレオチドなどを内因性リガンドとするP2受容体と共にプリン受容体と呼ばれる(表1、2)[1] [2]。
なお、本項における受容体の表記は、IUPHAR 国際薬理学連合でのデータベース掲載名に従った。
| 名称 | 下位分類 | リガンド | 作用機構 | |
| P1受容体 | A1、A2A、A2B,A3受容体 | アデノシン | Gタンパク質共役型受容体 | |
| P2受容体 | P2X受容体 | P2X1−7受容体 | ATP | リガンド依存性イオンチャンネル |
| P2Y受容体 | P2Y1, 2, 4, 6, 11−14受容体 | ヌクレオチド | Gタンパク質共役型受容体 | |
編集部にてWikipediaより翻訳、改変。
サブタイプ
A1受容体
Gi/Goタンパク質共役型受容体である。カフェインの作用標的としても知られている。脳や脊髄などの中枢神経系組織に加え、心臓や腎臓、肺といった末梢組織を含む全身の様々な組織に発現している[3] [4]。A1受容体はP2Y1受容体とヘテロマーを形成することも知られている[5]。これまでに数十種類の選択的作動薬や拮抗薬が合成され、その機能解明も進んでいる。例えば、脳において興奮性シナプス伝達を抑制的に制御していることが知られており[6]、A1受容体欠損マウスの海馬においては、シナプス伝達の異常興奮が観察される[7]。また、心筋の収縮力や心拍数の制御に加え、血管拡張作用、体温調節機構にも関与していると考えられている[7] [8]。A1受容体欠損マウスは、てんかん症状や痛覚過敏症状、不安関連行動を呈するほか、インスリン分泌の亢進や低酸素性障害や虚血ストレスに対する抵抗性の低下が観察される[7] [9]。
A2A受容体
Gsタンパク質共役型GPCRで、受容体刺激によりアデニル酸シクラーゼが活性化され、cAMPの産生が亢進する。A1受容体[10]や他の神経伝達物質受容体(ドーパミンD2受容体[11], D3受容体[12], 代謝型グルタミン酸受容体mGluR5[13], CB1受容体[14])とヘテロマーを形成することがある。A1受容体と同様に、カフェインが拮抗薬として作用する。また、A2A受容体に対する選択的作用薬としてCGS21680やDPMA、拮抗薬としてイストラデフィリンやSCH-58261がある[15]。A2A受容体は、生体内に幅広く分布し、線条体や海馬、冠血管、肺、血小板、腎臓などに発現する[16] [17] [18]。生理的役割としては、血管拡張、睡眠、神経活動制御があり、A2A受容体欠損マウスにおいて不安行動及び攻撃性増加や痛覚鈍麻、心拍数増加、血圧上昇、血小板凝集が見られる[19] [20]。最近、ミクログリア細胞にA2A受容体が発現し、突起の退縮に関与していることも報告された[21]。現在、A2A受容体作動薬レガデノソン(レキスキャン®、アステラス製薬/CVセラピューティクス)が心筋血流イメージングの薬物負荷剤として、A2A受容体拮抗薬イストラデフィリン(ノウリアスト®、協和発酵キリン)がパーキンソン病治療薬として用いられている。
A2B受容体
A2A受容体と同様にGs共役型GPCRであるが、アデノシンへの親和性が低く、アデノシンの他にネトリン-1の受容体としても機能する[22]。A2B受容体選択的作動薬としてBAY 60-6583やNECA、拮抗薬としてMRS-1754やMRE-2029-F20、CVT6883、PSB-1115などがある[15]。A2B受容体は、中枢神経系を含め、全身に広く発現しており[23]、特に脳や腸、腎臓、肺に多く、心臓や大動脈にもわずかに発現が見られるが、肝臓ではほとんど見られない[24]。また、肥満細胞や線維芽細胞にも発現する[23] [25]。A2B受容体欠損マウスにおいて、リポ多糖(LPS)による炎症及びサイトカイン産生の抑制やリンパ球の血管付着の増加[24]、肥満細胞の活性化及びIgE誘発アナフィラキシー亢進[25]、心筋虚血プレコンデショニングによる心臓保護作用の抑制が報告されている[26]。
A3受容体
薬理学的機能同定前にクローニングされた唯一のアデノシン受容体サブタイプで、Gi、GoあるいはGqタンパク質と共役し細胞内にシグナルを伝える[6]。肺や腎臓、心臓、脳、脾臓、肝臓など全身の様々な組織に発現しているが、その発現レベルは動物種間で大きく異なっている[4] [27]。例えば、ラットでは睾丸や肥満細胞で発現量が高いのに対して、ヒトでは肺や肝臓でその発現が高く、脳や大動脈での発現量は低い[3]。A3受容体欠損マウスにおいては、野生型マウスと比較して、眼圧の低下[28]や局所炎症反応の減少[29]など、いくつかの表現型の違いが観察される。
| 受容体 | 遺伝子 | Allen Brain Atlas | 細胞内情報伝達機構 | 作用 | アゴニスト | アンタゴニスト |
|---|---|---|---|---|---|---|
| A1受容体 | ADORA1 | 73496905 | Gi/o → cAMP↑/↓ | |||
| A2A受容体 | ADORA2A | 69855739 | Gs → cAMP↑ |
|
| |
| A2B受容体 | ADORA2B | 69855773 | Gs → cAMP↑ |
|
||
| A3受容体 | ADORA3 | 75828188 | Gi → cAMP↓ |
|
編集部にてWikipediaより翻訳、改変。
関連項目
参考文献
- ↑
Burnstock, G. (2007).
Physiology and pathophysiology of purinergic neurotransmission. Physiological reviews, 87(2), 659-797. [PubMed:17429044] [WorldCat] [DOI] - ↑
Burnstock, G. (2008).
Purinergic signalling and disorders of the central nervous system. Nature reviews. Drug discovery, 7(7), 575-90. [PubMed:18591979] [WorldCat] [DOI] - ↑ 3.0 3.1
Salvatore, C.A., Jacobson, M.A., Taylor, H.E., Linden, J., & Johnson, R.G. (1993).
Molecular cloning and characterization of the human A3 adenosine receptor. Proceedings of the National Academy of Sciences of the United States of America, 90(21), 10365-9. [PubMed:8234299] [PMC] [WorldCat] [DOI] - ↑ 4.0 4.1
Regard, J.B., Sato, I.T., & Coughlin, S.R. (2008).
Anatomical profiling of G protein-coupled receptor expression. Cell, 135(3), 561-71. [PubMed:18984166] [PMC] [WorldCat] [DOI] - ↑
Yoshioka, K., Hosoda, R., Kuroda, Y., & Nakata, H. (2002).
Hetero-oligomerization of adenosine A1 receptors with P2Y1 receptors in rat brains. FEBS letters, 531(2), 299-303. [PubMed:12417330] [WorldCat] [DOI] - ↑ 6.0 6.1
Dias, R.B., Rombo, D.M., Ribeiro, J.A., Henley, J.M., & Sebastião, A.M. (2013).
Adenosine: setting the stage for plasticity. Trends in neurosciences, 36(4), 248-57. [PubMed:23332692] [WorldCat] [DOI] - ↑ 7.0 7.1 7.2
Johansson, B., Halldner, L., Dunwiddie, T.V., Masino, S.A., Poelchen, W., Giménez-Llort, L., ..., & Fredholm, B.B. (2001).
Hyperalgesia, anxiety, and decreased hypoxic neuroprotection in mice lacking the adenosine A1 receptor. Proceedings of the National Academy of Sciences of the United States of America, 98(16), 9407-12. [PubMed:11470917] [PMC] [WorldCat] [DOI] - ↑
Brown, R., Ollerstam, A., Johansson, B., Skøtt, O., Gebre-Medhin, S., Fredholm, B., & Persson, A.E. (2001).
Abolished tubuloglomerular feedback and increased plasma renin in adenosine A1 receptor-deficient mice. American journal of physiology. Regulatory, integrative and comparative physiology, 281(5), R1362-7. [PubMed:11641103] [WorldCat] [DOI] - ↑
Wu, W.P., Hao, J.X., Halldner, L., Lövdahl, C., DeLander, G.E., Wiesenfeld-Hallin, Z., ..., & Xu, X.J. (2005).
Increased nociceptive response in mice lacking the adenosine A1 receptor. Pain, 113(3), 395-404. [PubMed:15661449] [WorldCat] [DOI] - ↑
Ciruela, F., Casadó, V., Rodrigues, R.J., Luján, R., Burgueño, J., Canals, M., ..., & Franco, R. (2006).
Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. The Journal of neuroscience : the official journal of the Society for Neuroscience, 26(7), 2080-7. [PubMed:16481441] [PMC] [WorldCat] [DOI] - ↑
Fuxe, K., Ferré, S., Canals, M., Torvinen, M., Terasmaa, A., Marcellino, D., ..., & Franco, R. (2005).
Adenosine A2A and dopamine D2 heteromeric receptor complexes and their function. Journal of molecular neuroscience : MN, 26(2-3), 209-20. [PubMed:16012194] [WorldCat] [DOI] - ↑
Torvinen, M., Marcellino, D., Canals, M., Agnati, L.F., Lluis, C., Franco, R., & Fuxe, K. (2005).
Adenosine A2A receptor and dopamine D3 receptor interactions: evidence of functional A2A/D3 heteromeric complexes. Molecular pharmacology, 67(2), 400-7. [PubMed:15539641] [WorldCat] [DOI] - ↑
Zezula, J., & Freissmuth, M. (2008).
The A(2A)-adenosine receptor: a GPCR with unique features? British journal of pharmacology, 153 Suppl 1, S184-90. [PubMed:18246094] [PMC] [WorldCat] [DOI] - ↑
Ferré, S., Goldberg, S.R., Lluis, C., & Franco, R. (2009).
Looking for the role of cannabinoid receptor heteromers in striatal function. Neuropharmacology, 56 Suppl 1, 226-34. [PubMed:18691604] [PMC] [WorldCat] [DOI] - ↑ 15.0 15.1
Jacobson, K.A., Balasubramanian, R., Deflorian, F., & Gao, Z.G. (2012).
G protein-coupled adenosine (P1) and P2Y receptors: ligand design and receptor interactions. Purinergic signalling, 8(3), 419-36. [PubMed:22371149] [PMC] [WorldCat] [DOI] - ↑
Rosin, D.L., Robeva, A., Woodard, R.L., Guyenet, P.G., & Linden, J. (1998).
Immunohistochemical localization of adenosine A2A receptors in the rat central nervous system. The Journal of comparative neurology, 401(2), 163-86. [PubMed:9822147] [WorldCat] - ↑
Peterfreund, R.A., MacCollin, M., Gusella, J., & Fink, J.S. (1996).
Characterization and expression of the human A2a adenosine receptor gene. Journal of neurochemistry, 66(1), 362-8. [PubMed:8522976] [WorldCat] [DOI] - ↑
Varani, K., Gessi, S., Dalpiaz, A., & Borea, P.A. (1996).
Pharmacological and biochemical characterization of purified A2a adenosine receptors in human platelet membranes by [3H]-CGS 21680 binding. British journal of pharmacology, 117(8), 1693-701. [PubMed:8732278] [PMC] [WorldCat] [DOI] - ↑
Satoh, S., Matsumura, H., Suzuki, F., & Hayaishi, O. (1996).
Promotion of sleep mediated by the A2a-adenosine receptor and possible involvement of this receptor in the sleep induced by prostaglandin D2 in rats. Proceedings of the National Academy of Sciences of the United States of America, 93(12), 5980-4. [PubMed:8650205] [PMC] [WorldCat] [DOI] - ↑
Ledent, C., Vaugeois, J.M., Schiffmann, S.N., Pedrazzini, T., El Yacoubi, M., Vanderhaeghen, J.J., ..., & Parmentier, M. (1997).
Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature, 388(6643), 674-8. [PubMed:9262401] [WorldCat] [DOI] - ↑
Orr, A.G., Orr, A.L., Li, X.J., Gross, R.E., & Traynelis, S.F. (2009).
Adenosine A(2A) receptor mediates microglial process retraction. Nature neuroscience, 12(7), 872-8. [PubMed:19525944] [PMC] [WorldCat] [DOI] - ↑
Corset, V., Nguyen-Ba-Charvet, K.T., Forcet, C., Moyse, E., Chédotal, A., & Mehlen, P. (2000).
Netrin-1-mediated axon outgrowth and cAMP production requires interaction with adenosine A2b receptor. Nature, 407(6805), 747-50. [PubMed:11048721] [WorldCat] [DOI] - ↑ 23.0 23.1
Feoktistov, I., & Biaggioni, I. (1997).
Adenosine A2B receptors. Pharmacological reviews, 49(4), 381-402. [PubMed:9443164] [WorldCat] - ↑ 24.0 24.1
Yang, D., Zhang, Y., Nguyen, H.G., Koupenova, M., Chauhan, A.K., Makitalo, M., ..., & Ravid, K. (2006).
The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. The Journal of clinical investigation, 116(7), 1913-23. [PubMed:16823489] [PMC] [WorldCat] [DOI] - ↑ 25.0 25.1
Hua, X., Kovarova, M., Chason, K.D., Nguyen, M., Koller, B.H., & Tilley, S.L. (2007).
Enhanced mast cell activation in mice deficient in the A2b adenosine receptor. The Journal of experimental medicine, 204(1), 117-28. [PubMed:17200408] [PMC] [WorldCat] [DOI] - ↑
Eckle, T., Krahn, T., Grenz, A., Köhler, D., Mittelbronn, M., Ledent, C., ..., & Eltzschig, H.K. (2007).
Cardioprotection by ecto-5'-nucleotidase (CD73) and A2B adenosine receptors. Circulation, 115(12), 1581-90. [PubMed:17353435] [WorldCat] [DOI] - ↑
Gessi, S., Merighi, S., Varani, K., Leung, E., Mac Lennan, S., & Borea, P.A. (2008).
The A3 adenosine receptor: an enigmatic player in cell biology. Pharmacology & therapeutics, 117(1), 123-40. [PubMed:18029023] [WorldCat] [DOI] - ↑
Avila, M.Y., Stone, R.A., & Civan, M.M. (2002).
Knockout of A3 adenosine receptors reduces mouse intraocular pressure. Investigative ophthalmology & visual science, 43(9), 3021-6. [PubMed:12202525] [WorldCat] - ↑
Wu, W.P., Hao, J.X., Halldner-Henriksson, L., Xu, X.J., Jacobson, M.A., Wiesenfeld-Hallin, Z., & Fredholm, B.B. (2002).
Decreased inflammatory pain due to reduced carrageenan-induced inflammation in mice lacking adenosine A3 receptors. Neuroscience, 114(3), 523-7. [PubMed:12220556] [WorldCat] [DOI]