「グリシン受容体」の版間の差分
細 →サブタイプ |
|||
| (2人の利用者による、間の15版が非表示) | |||
| 2行目: | 2行目: | ||
<font size="+1">[http://researchmap.jp/read0098270 赤木 宏行]</font><br> | <font size="+1">[http://researchmap.jp/read0098270 赤木 宏行]</font><br> | ||
''広島国際大学薬学部''<br> | ''広島国際大学薬学部''<br> | ||
DOI:<selfdoi /> 原稿受付日:2013年11月27日 原稿完成日:2013年12月28日<br> | |||
担当編集委員:[http://researchmap.jp/ | 担当編集委員:[http://researchmap.jp/2rikenbsi 林 康紀](独立行政法人理化学研究所 脳科学総合研究センター)<br> | ||
</div> | </div> | ||
{{box|text= | {{box|text= [[グリシン]]は、[[wikipedia:ja:脊椎動物|脊椎動物]]の神経系において[[神経伝達物質]]として機能している。[[ニューロン]]の[[シナプス]]膜に発現するグリシン受容体は、2個のαサブユニット、3個のβサブユニットから成る5量体構造であり、その中心部に陰イオンチャネルが形成される。さらに細胞質タンパク質の[[ゲフィリン]](gephyrin)が会合して、シナプス後膜に固定されている。イオンチャネルの内壁に面するαサブユニットの第2膜貫通領域には、グリシン受容体と同じく陰イオンチャネル型受容体である[[GABAA受容体|γ-アミノ酪酸(GABA)<sub>A</sub>受容体]]の同部位と共通のアミノ酸配列がみられ、この領域が陰イオン(生理的には主にCl<sup>-</sup>)の選択的透過性を担っているものと考えられる。グリシンのαサブユニットにはα<sub>1</sub>からα<sub>4</sub>までのサブファミリーが存在し、神経系におけるそれぞれのサブユニットの発現部位や発現の時期には違いが見られる。グリシン作動性シナプスの主要な機能は、[[興奮性シナプス]]伝達の抑制であり、[[wikipedia:ja:ほ乳類|ほ乳類]]では,主に[[脳幹]]や[[脊髄]]全域で作動している。近年、α<sub>3</sub>サブユニットを含むグリシン受容体が[[wikipedia:ja:炎症性疼痛|炎症性疼痛]]の制御に関与することが示され、注目されている。}} | ||
{{Pfam_box | |||
| Symbol = Neur_chan_memb | |||
| Name = Neurotransmitter-gated ion-channel transmembrane region | |||
| image = 2M6I.pdb | |||
| width = 250px | |||
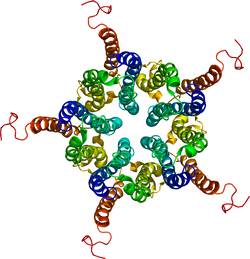
| caption = グリシン受容体チャネル。{{PDB2|2M6I}}による<ref><PubMed>23994010</pubmed></ref>。 | |||
| Pfam = PF02932 | |||
| InterPro = IPR006029 | |||
| SMART = | |||
| PROSITE = PDOC00209 | |||
| SCOP = 1cek | |||
| TCDB = 1.A.9 | |||
| OPM family = 14 | |||
| OPM protein = 2m6i | |||
}} | }} | ||
== | == グリシン受容体とは == | ||
グリシンはタンパク質を構成する重要な[[wikipedia:ja:α-アミノ酸|α-アミノ酸]]の一つであるが、それ自体が[[抑制性神経伝達物質]]して機能している。同様な作用を示す[[γ-アミノ酪酸]]([[GABA]])が[[中枢神経]]全体で広く作用するのに対し、グリシンは主に脳幹と脊髄で機能する。 | |||
グリシンが脊髄ニューロンに対し,抑制作用を引き起こすことは1950年代から知られていたが、1960年代後半になって、ネコの運動ニューロンで記録された[[抑制性シナプス]]後電位(IPSP)と、電気泳動的にそのニューロン近傍に投与されたグリシンによる[[膜電位]]変化が、共に[[wikipedia:ja:塩素イオン|塩素イオン]]([[wikipedia:ja:Cl-|Cl<sup>-</sup>]])の膜透過性を亢進させること、また、植物毒[[wikipedia:ja:ストリキニーネ|ストリキニーネ]]([[wikipedia:ja:ホミカ|ホミカ]]に含まれるインドールアルカロイドで、生体に強力な[[痙攣]]誘発作用を持つ)が両反応を抑制することが示され、グリシンの抑制性神経伝達物質としての役割が明確にされた<ref name=ref1>'''Aprison, M.H.'''<br>The discovery of the neurotransmitter role of glycine.<br> | |||
In:Glycine neurotransmission. (Ed. By Ottesen, O.P., and Strom-Mathisen, J.) <br>pp. 1-23 J''ohn Wiley, Chichester,'' 1990</ref>。 | In:Glycine neurotransmission. (Ed. By Ottesen, O.P., and Strom-Mathisen, J.) <br>pp. 1-23 J''ohn Wiley, Chichester,'' 1990</ref>。 | ||
グリシン受容体の分子実体を明らかにする研究は、まずストリキニーネがこの受容体に特異的に結合する性質を利用して、タンパク質を[[アフィニティーカラム]]で精製することから始められ、単離されたタンパク質の部分構造を基に、[[cDNA]]クローニングが行われ、複数のサブユニットの全長をコードするcDNAの塩基配列とそれらから推定されるアミノ酸配列が決定された<ref name=ref2><pubmed>3037383</pubmed></ref> <ref name=ref3><pubmed>2163264</pubmed></ref> <ref name=ref4><pubmed>1716350</pubmed></ref>。 | |||
== 構造 == | == 構造 == | ||
αサブユニットとβサブユニットから成る5量体を形成し、それ自体が陰イオン(主にCl- | αサブユニットとβサブユニットから成る5量体を形成し、それ自体が陰イオン(主にCl<sup>-</sup>)チャネルとして機能する[[リガンドゲート型イオンチャネル]]のファミリーに属する。さらに細胞内でゲフィリン(gephyrin)が[[裏打ちタンパク質]]として[[シナプス後膜]]に固定するアンカーの役割を果たしている<ref name=ref5><pubmed>7546736</pubmed></ref>。αサブユニットとβサブユニットの組成比(stoichiometry)は2:3であると考えられている<ref name=ref6><pubmed>15748848</pubmed></ref>。 | ||
αとβの両サブユニットは共に膜4回貫通型であり、第2膜貫通部位(TM2)がイオンチャネルの機能を担っている。αサブユニットのTM2にはTTVLTMTTという8個のアミノ酸配列が保存されており、この配列は、陰イオンチャネル内蔵型受容体であるGABA<sub>A</sub>受容体のすべてのサブユニットにも含まれていることから、この領域がCl―チャネルコアに相当するものと推定される。さらに、それを挟む形で配置されている[[アルギニン]](R)残基の陽性電荷がチャネルの陰イオン選択性に関与していると考えられる<ref name=ref2 /> <ref name=ref4 />。 | |||
グリシン受容体を構成するα、βサブユニットを[[培養細胞]]に人工的に発現させて、機能解析をした結果から、αサブユニット上にアゴニストであるグリシンや[[wikipedia:ja:β-アラニン|β-アラニン]]、アンタゴニストであるストリキニーネが結合する領域が存在することは確かである<ref name=ref7><pubmed>1281418</pubmed></ref> <ref name=ref8><pubmed>16805771</pubmed></ref>が、アゴニストの結合部位が隣接するα-βサブユニットに跨がっているという報告もある<ref name=ref6 />。GABAA受容体では、アゴニスト結合能を持たないサブユニット上に、チャネルの機能調節を行う[[ベンゾジアゼピン]]系薬物や[[バルビツール酸]]系の薬物が結合する部位が存在するが、グリシン受容体では、そのような薬物の調節様式は今のところ明らかにされていない。[[wikipedia:ja:ハロタン|ハロタン]]や[[wikipedia:ja:エンフルレン|エンフルレン]]などの[[揮発性麻酔薬]]は、GABA<sub>A</sub>受容体とグリシン受容体のチャネル領域に作用して機能を増強させる<ref name=ref9><pubmed>1667416</pubmed></ref>。 | |||
== サブタイプ == | == サブタイプ == | ||
αサブユニットにはα<sub>1</sub>からα<sub>4</sub>の4種類のサブタイプが存在する<ref name=ref10><pubmed>2176214</pubmed></ref>。これらのサブタイプはアミノ酸配列において71%-83%の相同性を持ち、特に4箇所の細胞膜貫通領域は類似性が高い(90%以上の相同性)。α<sub>1</sub>-α<sub>4</sub> | αサブユニットにはα<sub>1</sub>からα<sub>4</sub>の4種類のサブタイプが存在する<ref name=ref10><pubmed>2176214</pubmed></ref>。これらのサブタイプはアミノ酸配列において71%-83%の相同性を持ち、特に4箇所の細胞膜貫通領域は類似性が高い(90%以上の相同性)。α<sub>1</sub>-α<sub>4</sub>サブタイプをコードする[[mRNA]]を[[アフリカツメガエル]]の[[wikipedia:ja:卵母細胞|卵母細胞]]に注入して発現させたホモメリック受容体は、いずれもグリシンに応答するCl<sup>-</sup>チャネル機能を示すが、部分アゴニストであるβ-アラニンや、アンタゴニストであるストリキニーネに対する感受性は異なる<ref name=ref11><pubmed>1707830</pubmed></ref>。βサブユニットやゲフェィンのサブタイプの存在は今のところ明らかにされていない。 | ||
== 発現(組織分布) == | == 発現(組織分布) == | ||
[[In situ hybridization法]]により、グリシン受容体の各サブユニットをコードするmRNAの分布が調べられている<ref name=ref12><pubmed>1651228</pubmed></ref> <ref name=ref13><pubmed>1330217</pubmed></ref> <ref name=ref14><pubmed>8602277</pubmed></ref>。 | |||
;α<sub>1</sub>サブユニット | |||
:成熟動物の脊髄、[[延髄]]に豊富に発現している。[[脊髄前角]]の[[運動ニューロン]]プールは強く標識されるが、[[後角]]の細胞群も中程度のラベルが多数見られることから、脊髄[[灰白質]]のほぼ全域に発現していると思われる。[[三叉神経]][[脊髄路核]]、[[蝸牛神経]]核を含む下部脳幹でも発現している。[[大脳皮質]]など上部脳での発現は観察されない。成長期[[ラット]]脊髄では、胎生19日ごろから発現が始まり、以後、生後発達とともに発現量が増加する。 | |||
;α<sub>2</sub>サブユニット | |||
:胎児期、生後の幼若期の脳、脊髄に広く発現している。胎生14日の時期にはすでに[[終脳]]、[[間脳]]、[[中脳]]、脊髄で強いシグナルが観察される。動物の日令とともにα<sub>2</sub>のシグナルは減少し、成熟期には消失するが、[[海馬]]を含む一部の大脳皮質では、成熟期においても発現している<ref name=ref14 />。 | |||
;α<sub>3</sub>サブユニット | |||
:成熟期動物の脊髄、[[大脳辺縁系]]、[[小脳顆粒細胞]]層に発現しているが、量的割合はα<sub>1</sub>と比べて低い。脊髄では後角表層(laminae I, II)に限局して発現している<ref name=ref8 />。 | |||
;α<sub>4</sub>サブユニット | |||
:胎児期の脊髄に発現が観られるが、発現量は著しく少ない。 | |||
;βサブユニット | |||
:α<sub>1</sub>サブユニットと同様に胎児期・幼若期の脳脊髄には少なく、生後発達とともに発現量が増加するパターンを示す。 | |||
== 機能 == | == 機能 == | ||
===興奮抑制機能=== | |||
グリシン受容体の重要な役割は、脊髄における興奮性シナプス伝達の抑制である。グリシン受容体チャネルを介して透過するCl<sup>-</sup>の平衡電位は、通常、[[静止膜電位]]より[[過分極側]]であるため、IPSPが生じている間はニューロンの興奮性が低下する。 | |||
ただし、陰イオンチャネル型のグリシン受容体も幼若期には運動ニューロンに脱分極反応を引き起こす<ref name=ref18>'''大塚正徳'''<br>化学伝達物質の歴史と現状(大塚正徳、竹内昭偏)<br>''伝達物質と受容物質'' pp.1-18 産業図書、1976</ref>。同様の現象は、GABA<sub>A</sub>受容体を介する反応でも生じる。これは、両者の受容体のイオンチャネルを透過するCl<sup>-</sup>の細胞内濃度が成熟期のニューロンよりも高いためと考えられる。ある種のニューロンの細胞内Cl<sup>-</sup>濃度は, [[K+,Cl-共輸送体|K<sup>+</sup>,Cl<sup>-</sup>共輸送体]](KCC2)により調節されており、このトランスポーターは生後に発現する<ref name=ref19><pubmed>9930699</pubmed></ref>。従ってグリシンやGABA<sub>A</sub>受容体を介する反応は、動物の成熟が進むにつれて脱分極性から過分極性に変化する。このような経時的変化は、神経回路やシナプスの可塑的変化と密接な関係があると考えられている。 | |||
[[file:higashijima_fig1.png|thumb|300px| '''図1. レンショウ細胞'''<br>文献<ref name=Kandel>'''Eric R. Kandel, James H. Schwartz and Thomas M. Jessell'''<br>Principles of Neural Science, 4th edition<br>''McGraw-Hill Companies'', 2000</ref>より改変]] | |||
===グリシン性シナプスの分布=== | |||
最も古くから知られている神経回路は脊髄前角の[[脊髄介在ニューロン|介在ニューロン]]([[レンショウ細胞]])を介する運動ニューロンの興奮抑制([[レンショウ抑制]])である<ref name=ref15><pubmed>13222354</pubmed></ref>。運動ニューロン細胞体の興奮により軸索に発生した活動電位が、軸索側枝を介してレンショウ細胞の興奮を促し、その神経終末と運動ニューロンとのシナプスで生じるIPSPが続発する運動ニューロンの興奮を抑制する[[反回性回路]]が存在し、グリシンはレンショウ細胞の終末から放出される伝達物質として働く。運動ニューロン膜のグリシン作動性IPSPは、運動ニューロンに限らず、脊髄後角のニューロンからも記録されている。ストリキニーネは、脊髄のグリシン作動性抑制シナプス伝達を遮断することで、運動ニューロンの異常興奮を引き起こし、四肢の骨格筋の[[強直性痙攣]]や[[wikipedia:ja:横隔膜筋|横隔膜筋]]の痙攣による呼吸麻痺を引き起こす<ref name=ref1 />。 | |||
脊髄では、胎児・幼若期に豊富なグリシン受容体α<sub>2</sub>サブユニットから成熟期に多いα<sub>1</sub>サブユニットへの発現の切り替えが起こる(発現の項参照)。α<sub>2</sub>サブユニットから成るホモメリックグリシン受容体の単一チャネル[[開口時間]]は、α<sub>1</sub>ホモメリック受容体のそれと比べて数十倍長い。ラットの脊髄ニューロンでは、グリシン受容体単一チャネルの開口時間も成長にともなって短くなる。これと平行して、ラット脊髄スライス標本で記録されるグリシン作動性シナプス電流の減衰時間経過が日令とともに短くなる現象が示されている<ref name=ref7 />。これらのことは、グリシン受容体の構成サブユニットがα<sub>2</sub>優位型からα<sub>1</sub>優位型に置き換わることにより生じる可能性を示唆している。成長に伴う運動機能の発達に応じて、それを制御するグリシン作動性シナプスの可塑性が生じているものと考えられる。これに関して、α<sub>2</sub>サブユニットを[[ノックアウト]]したマウスの発育は妨げられないという報告があるので、発達時期のグリシン作動性シナプスの可塑性には、多様な仕組みが関与しているのかもしれない<ref name=ref16><pubmed>16847326</pubmed></ref>。 | |||
α<sub>3</sub>サブユニットの発現量はα<sub>1</sub>サブユニットのそれと比べて低いが、脊髄においては、α<sub>1</sub>が灰白質全体に発現するのに対し、α<sub>3</sub>は後角の表層(laminae I, II)に限局して発現する。この部位では、[[一次知覚ニューロン]]のうち、[[痛覚]]などの侵害性感覚に関与する[[C線維]]がシナプス形成をすることが知られている。生体内炎症性発痛物質のひとつである[[プロスタグランジン]](PG)E2が、脊髄においてα<sub>3</sub>サブユニットを含むグリシン受容体チャネルをリン酸化して機能低下を引き起こすことが報告されており、炎症性疼痛の制御にα<sub>3</sub>グリシン受容体が関与する可能性が示唆されている<ref name=ref17><pubmed>15131310</pubmed></ref>。 | |||
α | α<sub>4</sub>サブユニットの機能的役割は、今のところ明らかにされていない。 | ||
== 疾患との関わり == | |||
グリシン受容体の異常に起因する先天性疾患が、[[マウス]]、[[ウシ]]、ヒトや[[ゼブラフィッシュ]]で知られている<ref name=ref20><pubmed>2845573</pubmed></ref> <ref name=ref21><pubmed>7946325</pubmed></ref> <ref name=ref22><pubmed>8298642</pubmed></ref> <ref name=ref23><pubmed>15928085</pubmed></ref>。 | |||
ヒトの場合、[[hyperekplexia]](別名[[びっくり病]]、[[startle disease]])がそれに該当する<ref name=ref22 />。この疾患は、出生後に発症し、音などの外界刺激に過剰反応して四肢の運動異常が続く。この病気の患者遺伝子解析が行われた結果、α<sub>1</sub>サブユニットの271番目のアミノ酸が野生型のRからL/Qに変異していることが判明した。271R残基は、TM2膜貫通部に隣接する細胞外領域にあり、陰イオン選択性を決める重要なアミノ酸のひとつである。R残基が持つ陽性電荷が失われることで、グリシン受容体が機能しなくなるものと推定される。 | |||
== 関連項目 == | == 関連項目 == | ||
*[[抑制性アミノ酸]] | |||
*[[グリシン]] | |||
*[[脊髄介在ニューロン]] | |||
*[[シナプス伝達]] | *[[シナプス伝達]] | ||
*[[神経伝達物質受容体]] | *[[神経伝達物質受容体]] | ||
*[[イオンチャネル型受容体]] | *[[イオンチャネル型受容体]] | ||
*[[GABAA受容体 | *[[GABAA受容体|GABA<sub>A</sub>受容体]] | ||
== 参考文献 == | == 参考文献 == | ||
<references /> | <references /> | ||
2021年12月15日 (水) 20:48時点における最新版
赤木 宏行
広島国際大学薬学部
DOI:10.14931/bsd.3921 原稿受付日:2013年11月27日 原稿完成日:2013年12月28日
担当編集委員:林 康紀(独立行政法人理化学研究所 脳科学総合研究センター)
グリシンは、脊椎動物の神経系において神経伝達物質として機能している。ニューロンのシナプス膜に発現するグリシン受容体は、2個のαサブユニット、3個のβサブユニットから成る5量体構造であり、その中心部に陰イオンチャネルが形成される。さらに細胞質タンパク質のゲフィリン(gephyrin)が会合して、シナプス後膜に固定されている。イオンチャネルの内壁に面するαサブユニットの第2膜貫通領域には、グリシン受容体と同じく陰イオンチャネル型受容体であるγ-アミノ酪酸(GABA)A受容体の同部位と共通のアミノ酸配列がみられ、この領域が陰イオン(生理的には主にCl-)の選択的透過性を担っているものと考えられる。グリシンのαサブユニットにはα1からα4までのサブファミリーが存在し、神経系におけるそれぞれのサブユニットの発現部位や発現の時期には違いが見られる。グリシン作動性シナプスの主要な機能は、興奮性シナプス伝達の抑制であり、ほ乳類では,主に脳幹や脊髄全域で作動している。近年、α3サブユニットを含むグリシン受容体が炎症性疼痛の制御に関与することが示され、注目されている。
| Neurotransmitter-gated ion-channel transmembrane region | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | Neur_chan_memb | ||||||||
| Pfam | PF02932 | ||||||||
| InterPro | IPR006029 | ||||||||
| PROSITE | PDOC00209 | ||||||||
| SCOP | 1cek | ||||||||
| SUPERFAMILY | 1cek | ||||||||
| TCDB | 1.A.9 | ||||||||
| OPM superfamily | 14 | ||||||||
| OPM protein | 2m6i | ||||||||
| |||||||||
グリシン受容体とは
グリシンはタンパク質を構成する重要なα-アミノ酸の一つであるが、それ自体が抑制性神経伝達物質して機能している。同様な作用を示すγ-アミノ酪酸(GABA)が中枢神経全体で広く作用するのに対し、グリシンは主に脳幹と脊髄で機能する。
グリシンが脊髄ニューロンに対し,抑制作用を引き起こすことは1950年代から知られていたが、1960年代後半になって、ネコの運動ニューロンで記録された抑制性シナプス後電位(IPSP)と、電気泳動的にそのニューロン近傍に投与されたグリシンによる膜電位変化が、共に塩素イオン(Cl-)の膜透過性を亢進させること、また、植物毒ストリキニーネ(ホミカに含まれるインドールアルカロイドで、生体に強力な痙攣誘発作用を持つ)が両反応を抑制することが示され、グリシンの抑制性神経伝達物質としての役割が明確にされた[2]。
グリシン受容体の分子実体を明らかにする研究は、まずストリキニーネがこの受容体に特異的に結合する性質を利用して、タンパク質をアフィニティーカラムで精製することから始められ、単離されたタンパク質の部分構造を基に、cDNAクローニングが行われ、複数のサブユニットの全長をコードするcDNAの塩基配列とそれらから推定されるアミノ酸配列が決定された[3] [4] [5]。
構造
αサブユニットとβサブユニットから成る5量体を形成し、それ自体が陰イオン(主にCl-)チャネルとして機能するリガンドゲート型イオンチャネルのファミリーに属する。さらに細胞内でゲフィリン(gephyrin)が裏打ちタンパク質としてシナプス後膜に固定するアンカーの役割を果たしている[6]。αサブユニットとβサブユニットの組成比(stoichiometry)は2:3であると考えられている[7]。
αとβの両サブユニットは共に膜4回貫通型であり、第2膜貫通部位(TM2)がイオンチャネルの機能を担っている。αサブユニットのTM2にはTTVLTMTTという8個のアミノ酸配列が保存されており、この配列は、陰イオンチャネル内蔵型受容体であるGABAA受容体のすべてのサブユニットにも含まれていることから、この領域がCl―チャネルコアに相当するものと推定される。さらに、それを挟む形で配置されているアルギニン(R)残基の陽性電荷がチャネルの陰イオン選択性に関与していると考えられる[3] [5]。
グリシン受容体を構成するα、βサブユニットを培養細胞に人工的に発現させて、機能解析をした結果から、αサブユニット上にアゴニストであるグリシンやβ-アラニン、アンタゴニストであるストリキニーネが結合する領域が存在することは確かである[8] [9]が、アゴニストの結合部位が隣接するα-βサブユニットに跨がっているという報告もある[7]。GABAA受容体では、アゴニスト結合能を持たないサブユニット上に、チャネルの機能調節を行うベンゾジアゼピン系薬物やバルビツール酸系の薬物が結合する部位が存在するが、グリシン受容体では、そのような薬物の調節様式は今のところ明らかにされていない。ハロタンやエンフルレンなどの揮発性麻酔薬は、GABAA受容体とグリシン受容体のチャネル領域に作用して機能を増強させる[10]。
サブタイプ
αサブユニットにはα1からα4の4種類のサブタイプが存在する[11]。これらのサブタイプはアミノ酸配列において71%-83%の相同性を持ち、特に4箇所の細胞膜貫通領域は類似性が高い(90%以上の相同性)。α1-α4サブタイプをコードするmRNAをアフリカツメガエルの卵母細胞に注入して発現させたホモメリック受容体は、いずれもグリシンに応答するCl-チャネル機能を示すが、部分アゴニストであるβ-アラニンや、アンタゴニストであるストリキニーネに対する感受性は異なる[12]。βサブユニットやゲフェィンのサブタイプの存在は今のところ明らかにされていない。
発現(組織分布)
In situ hybridization法により、グリシン受容体の各サブユニットをコードするmRNAの分布が調べられている[13] [14] [15]。
- α1サブユニット
- 成熟動物の脊髄、延髄に豊富に発現している。脊髄前角の運動ニューロンプールは強く標識されるが、後角の細胞群も中程度のラベルが多数見られることから、脊髄灰白質のほぼ全域に発現していると思われる。三叉神経脊髄路核、蝸牛神経核を含む下部脳幹でも発現している。大脳皮質など上部脳での発現は観察されない。成長期ラット脊髄では、胎生19日ごろから発現が始まり、以後、生後発達とともに発現量が増加する。
- α2サブユニット
- 胎児期、生後の幼若期の脳、脊髄に広く発現している。胎生14日の時期にはすでに終脳、間脳、中脳、脊髄で強いシグナルが観察される。動物の日令とともにα2のシグナルは減少し、成熟期には消失するが、海馬を含む一部の大脳皮質では、成熟期においても発現している[15]。
- α4サブユニット
- 胎児期の脊髄に発現が観られるが、発現量は著しく少ない。
- βサブユニット
- α1サブユニットと同様に胎児期・幼若期の脳脊髄には少なく、生後発達とともに発現量が増加するパターンを示す。
機能
興奮抑制機能
グリシン受容体の重要な役割は、脊髄における興奮性シナプス伝達の抑制である。グリシン受容体チャネルを介して透過するCl-の平衡電位は、通常、静止膜電位より過分極側であるため、IPSPが生じている間はニューロンの興奮性が低下する。
ただし、陰イオンチャネル型のグリシン受容体も幼若期には運動ニューロンに脱分極反応を引き起こす[16]。同様の現象は、GABAA受容体を介する反応でも生じる。これは、両者の受容体のイオンチャネルを透過するCl-の細胞内濃度が成熟期のニューロンよりも高いためと考えられる。ある種のニューロンの細胞内Cl-濃度は, K+,Cl-共輸送体(KCC2)により調節されており、このトランスポーターは生後に発現する[17]。従ってグリシンやGABAA受容体を介する反応は、動物の成熟が進むにつれて脱分極性から過分極性に変化する。このような経時的変化は、神経回路やシナプスの可塑的変化と密接な関係があると考えられている。

文献[18]より改変
グリシン性シナプスの分布
最も古くから知られている神経回路は脊髄前角の介在ニューロン(レンショウ細胞)を介する運動ニューロンの興奮抑制(レンショウ抑制)である[19]。運動ニューロン細胞体の興奮により軸索に発生した活動電位が、軸索側枝を介してレンショウ細胞の興奮を促し、その神経終末と運動ニューロンとのシナプスで生じるIPSPが続発する運動ニューロンの興奮を抑制する反回性回路が存在し、グリシンはレンショウ細胞の終末から放出される伝達物質として働く。運動ニューロン膜のグリシン作動性IPSPは、運動ニューロンに限らず、脊髄後角のニューロンからも記録されている。ストリキニーネは、脊髄のグリシン作動性抑制シナプス伝達を遮断することで、運動ニューロンの異常興奮を引き起こし、四肢の骨格筋の強直性痙攣や横隔膜筋の痙攣による呼吸麻痺を引き起こす[2]。
脊髄では、胎児・幼若期に豊富なグリシン受容体α2サブユニットから成熟期に多いα1サブユニットへの発現の切り替えが起こる(発現の項参照)。α2サブユニットから成るホモメリックグリシン受容体の単一チャネル開口時間は、α1ホモメリック受容体のそれと比べて数十倍長い。ラットの脊髄ニューロンでは、グリシン受容体単一チャネルの開口時間も成長にともなって短くなる。これと平行して、ラット脊髄スライス標本で記録されるグリシン作動性シナプス電流の減衰時間経過が日令とともに短くなる現象が示されている[8]。これらのことは、グリシン受容体の構成サブユニットがα2優位型からα1優位型に置き換わることにより生じる可能性を示唆している。成長に伴う運動機能の発達に応じて、それを制御するグリシン作動性シナプスの可塑性が生じているものと考えられる。これに関して、α2サブユニットをノックアウトしたマウスの発育は妨げられないという報告があるので、発達時期のグリシン作動性シナプスの可塑性には、多様な仕組みが関与しているのかもしれない[20]。
α3サブユニットの発現量はα1サブユニットのそれと比べて低いが、脊髄においては、α1が灰白質全体に発現するのに対し、α3は後角の表層(laminae I, II)に限局して発現する。この部位では、一次知覚ニューロンのうち、痛覚などの侵害性感覚に関与するC線維がシナプス形成をすることが知られている。生体内炎症性発痛物質のひとつであるプロスタグランジン(PG)E2が、脊髄においてα3サブユニットを含むグリシン受容体チャネルをリン酸化して機能低下を引き起こすことが報告されており、炎症性疼痛の制御にα3グリシン受容体が関与する可能性が示唆されている[21]。
α4サブユニットの機能的役割は、今のところ明らかにされていない。
疾患との関わり
グリシン受容体の異常に起因する先天性疾患が、マウス、ウシ、ヒトやゼブラフィッシュで知られている[22] [23] [24] [25]。
ヒトの場合、hyperekplexia(別名びっくり病、startle disease)がそれに該当する[24]。この疾患は、出生後に発症し、音などの外界刺激に過剰反応して四肢の運動異常が続く。この病気の患者遺伝子解析が行われた結果、α1サブユニットの271番目のアミノ酸が野生型のRからL/Qに変異していることが判明した。271R残基は、TM2膜貫通部に隣接する細胞外領域にあり、陰イオン選択性を決める重要なアミノ酸のひとつである。R残基が持つ陽性電荷が失われることで、グリシン受容体が機能しなくなるものと推定される。
関連項目
参考文献
- ↑
Mowrey, D.D., Cui, T., Jia, Y., Ma, D., Makhov, A.M., Zhang, P., ..., & Xu, Y. (2013).
Open-channel structures of the human glycine receptor α1 full-length transmembrane domain. Structure (London, England : 1993), 21(10), 1897-904. [PubMed:23994010] [PMC] [WorldCat] [DOI] - ↑ 2.0 2.1 Aprison, M.H.
The discovery of the neurotransmitter role of glycine.
In:Glycine neurotransmission. (Ed. By Ottesen, O.P., and Strom-Mathisen, J.)
pp. 1-23 John Wiley, Chichester, 1990 - ↑ 3.0 3.1
Grenningloh, G., Rienitz, A., Schmitt, B., Methfessel, C., Zensen, M., Beyreuther, K., ..., & Betz, H. (1987).
The strychnine-binding subunit of the glycine receptor shows homology with nicotinic acetylcholine receptors. Nature, 328(6127), 215-20. [PubMed:3037383] [WorldCat] [DOI] - ↑
Grenningloh, G., Pribilla, I., Prior, P., Multhaup, G., Beyreuther, K., Taleb, O., & Betz, H. (1990).
Cloning and expression of the 58 kd beta subunit of the inhibitory glycine receptor. Neuron, 4(6), 963-70. [PubMed:2163264] [WorldCat] [DOI] - ↑ 5.0 5.1
Akagi, H., Hirai, K., & Hishinuma, F. (1991).
Functional properties of strychnine-sensitive glycine receptors expressed in Xenopus oocytes injected with a single mRNA. Neuroscience research, 11(1), 28-40. [PubMed:1716350] [WorldCat] [DOI] - ↑
Meyer, G., Kirsch, J., Betz, H., & Langosch, D. (1995).
Identification of a gephyrin binding motif on the glycine receptor beta subunit. Neuron, 15(3), 563-72. [PubMed:7546736] [WorldCat] [DOI] - ↑ 7.0 7.1
Grudzinska, J., Schemm, R., Haeger, S., Nicke, A., Schmalzing, G., Betz, H., & Laube, B. (2005).
The beta subunit determines the ligand binding properties of synaptic glycine receptors. Neuron, 45(5), 727-39. [PubMed:15748848] [WorldCat] [DOI] - ↑ 8.0 8.1
Takahashi, T., Momiyama, A., Hirai, K., Hishinuma, F., & Akagi, H. (1992).
Functional correlation of fetal and adult forms of glycine receptors with developmental changes in inhibitory synaptic receptor channels. Neuron, 9(6), 1155-61. [PubMed:1281418] [WorldCat] [DOI] - ↑ 9.0 9.1
Betz, H., & Laube, B. (2006).
Glycine receptors: recent insights into their structural organization and functional diversity. Journal of neurochemistry, 97(6), 1600-10. [PubMed:16805771] [WorldCat] [DOI] - ↑
Wakamori, M., Ikemoto, Y., & Akaike, N. (1991).
Effects of two volatile anesthetics and a volatile convulsant on the excitatory and inhibitory amino acid responses in dissociated CNS neurons of the rat. Journal of neurophysiology, 66(6), 2014-21. [PubMed:1667416] [WorldCat] [DOI] - ↑
Kuhse, J., Schmieden, V., & Betz, H. (1990).
Identification and functional expression of a novel ligand binding subunit of the inhibitory glycine receptor. The Journal of biological chemistry, 265(36), 22317-20. [PubMed:2176214] [WorldCat] - ↑
Akagi, H., Hirai, K., & Hishinuma, F. (1991).
Cloning of a glycine receptor subtype expressed in rat brain and spinal cord during a specific period of neuronal development. FEBS letters, 281(1-2), 160-6. [PubMed:1707830] [WorldCat] [DOI] - ↑
Malosio, M.L., Marquèze-Pouey, B., Kuhse, J., & Betz, H. (1991).
Widespread expression of glycine receptor subunit mRNAs in the adult and developing rat brain. The EMBO journal, 10(9), 2401-9. [PubMed:1651228] [PMC] [WorldCat] - ↑
Sato, K., Kiyama, H., & Tohyama, M. (1992).
Regional distribution of cells expressing glycine receptor alpha 2 subunit mRNA in the rat brain. Brain research, 590(1-2), 95-108. [PubMed:1330217] [WorldCat] [DOI] - ↑ 15.0 15.1
Watanabe, E., & Akagi, H. (1995).
Distribution patterns of mRNAs encoding glycine receptor channels in the developing rat spinal cord. Neuroscience research, 23(4), 377-82. [PubMed:8602277] [WorldCat] [DOI] - ↑ 大塚正徳
化学伝達物質の歴史と現状(大塚正徳、竹内昭偏)
伝達物質と受容物質 pp.1-18 産業図書、1976 - ↑
Rivera, C., Voipio, J., Payne, J.A., Ruusuvuori, E., Lahtinen, H., Lamsa, K., ..., & Kaila, K. (1999).
The K+/Cl- co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature, 397(6716), 251-5. [PubMed:9930699] [WorldCat] [DOI] - ↑ Eric R. Kandel, James H. Schwartz and Thomas M. Jessell
Principles of Neural Science, 4th edition
McGraw-Hill Companies, 2000 - ↑
ECCLES, J.C., FATT, P., & KOKETSU, K. (1954).
Cholinergic and inhibitory synapses in a pathway from motor-axon collaterals to motoneurones. The Journal of physiology, 126(3), 524-62. [PubMed:13222354] [PMC] [WorldCat] [DOI] - ↑
Young-Pearse, T.L., Ivic, L., Kriegstein, A.R., & Cepko, C.L. (2006).
Characterization of mice with targeted deletion of glycine receptor alpha 2. Molecular and cellular biology, 26(15), 5728-34. [PubMed:16847326] [PMC] [WorldCat] [DOI] - ↑
Harvey, R.J., Depner, U.B., Wässle, H., Ahmadi, S., Heindl, C., Reinold, H., ..., & Müller, U. (2004).
GlyR alpha3: an essential target for spinal PGE2-mediated inflammatory pain sensitization. Science (New York, N.Y.), 304(5672), 884-7. [PubMed:15131310] [WorldCat] [DOI] - ↑
Gundlach, A.L., Dodd, P.R., Grabara, C.S., Watson, W.E., Johnston, G.A., Harper, P.A., ..., & Healy, P.J. (1988).
Deficit of spinal cord glycine/strychnine receptors in inherited myoclonus of Poll Hereford calves. Science (New York, N.Y.), 241(4874), 1807-10. [PubMed:2845573] [WorldCat] [DOI] - ↑
Mülhardt, C., Fischer, M., Gass, P., Simon-Chazottes, D., Guénet, J.L., Kuhse, J., ..., & Becker, C.M. (1994).
The spastic mouse: aberrant splicing of glycine receptor beta subunit mRNA caused by intronic insertion of L1 element. Neuron, 13(4), 1003-15. [PubMed:7946325] [WorldCat] [DOI] - ↑ 24.0 24.1
Shiang, R., Ryan, S.G., Zhu, Y.Z., Hahn, A.F., O'Connell, P., & Wasmuth, J.J. (1993).
Mutations in the alpha 1 subunit of the inhibitory glycine receptor cause the dominant neurologic disorder, hyperekplexia. Nature genetics, 5(4), 351-8. [PubMed:8298642] [WorldCat] [DOI] - ↑
Hirata, H., Saint-Amant, L., Downes, G.B., Cui, W.W., Zhou, W., Granato, M., & Kuwada, J.Y. (2005).
Zebrafish bandoneon mutants display behavioral defects due to a mutation in the glycine receptor beta-subunit. Proceedings of the National Academy of Sciences of the United States of America, 102(23), 8345-50. [PubMed:15928085] [PMC] [WorldCat] [DOI]