フォリスタチン
上田 洋司、土田 邦博
藤田医科大学 医科学研究センター 難病治療学部門
DOI:10.14931/bsd.10938 原稿受付日:2025年3月25日 原稿完成日:2025年4月4日
担当編集委員:山形 方人(ハーバード大学・脳科学センター)
英: follistatin
フォリスタチンは、下垂体前葉からの卵胞刺激ホルモン(FSH)分泌を抑制する糖タンパク質として発見された分子量31〜39 kDaの一本鎖構造を持つタンパク質である。スプライシングによりFS315とFS288の2つのアイソフォームが存在し、さらに分解産物であるFS303も生体内に確認されている。アクチビンと高親和性で結合し、アクチビンの受容体への結合を阻害してシグナル伝達を遮断することで、アクチビンの作用を抑制する。また、マイオスタチンや成長分化因子11 (growth differentiation factor 11; GDF11)、さらには一部の骨形成因子(BMP)にも結合し、これらの阻害因子としても機能することが分かっている。一方で、インヒビンにも弱く結合するが、その生理活性を阻害する作用はない。フォリスタチンは特徴的なフォリスタチンドメインを持ち、TGF-βファミリーとは異なる構造を有するため、両者を区別する必要がある。
フォリスタチンとは
ブタの卵胞液からインヒビンを精製する過程で、インヒビンとは異なった分画に下垂体前葉培養系でFSHの分泌を抑制する分子量31〜39kDaのシステインに富んだ一本鎖糖タンパク質が発見され、フォリスタチンと命名された[1]。FSHの分泌を抑制する作用はインヒビンと類似している。その後の解析から、アクチビンに高親和性 (Kd:500 pM程度)で結合しその作用を阻害することがわかり、その機能が解明された [2]。トランスフォーミング成長因子β (transforming growth factor-β; TGF-β)ファミリー分子に細胞外分泌型の阻害結合分子があることが最初にわかった例である。TGF-βファミリー分子である骨形成タンパク質 (bone morphogenetic protein; BMP)の機能を阻害するコーディン、ノギンなどのモデルケースとなった。フォリスタチンは、卵巣や卵胞液でアクチビンと複合体を形成しているが、フォリスタチンから遊離したアクチビンは生理活性を示すようになる。フォリスタチンは、アクチビンA、B、ABとの結合と阻害活性が強力であるが、次に親和性が高いのがマイオスタチン、成長分化因子11 (growth differentiation factor-11; GDF11)であり、フォリスタチンはこれらの分子も阻害する。

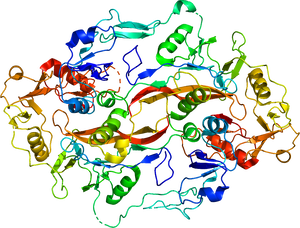
内部のアクチビン(シアン、青)をフォリスタチンが包み込む構造を取る。PDB 2B0U。

B. フォリスタチンによるアクチビンのシグナル伝達の阻害の模式図
中央がアクチビン(赤)
構造
フォリスタチン遺伝子はヒトを含めて種間でよく保存されており、6個のエクソンで構成されている。6番目のエクソンのスプライシングの違いにより、FS315とFS288の2つのバリアントが産生される。さらに分解産物であるFS303も生体内に確認されている(図1)。FS288は細胞との結合性が高く、アクチビン結合能と阻害活性がFS315より強い [3][4]。
フォリスタチンタンパク質は分子内にN末端領域(FSN)とシステインに富んだ3個のフォリスタチンドメイン (FSD1-3)を持つ糖付加ポリペプチドである。各FSDは10個のシステインを含んでおり、Kazal型のプロテアーゼインヒビターと構造上の類似性が見られるが、その活性は検出されない。
アクチビンとの結合と阻害活性には全体の分子構造が重要であるが、FSD1、FSD2が特に重要である。アクチビンと結合していないフリーの状態では、カルボキシル末端の酸性領域とFSD1の塩基性領域/ヘパリン結合領域とが相互作用しFS288よりもコンパクトな高次構造を取ると考えられている [5]。アクチビンと結合すると、その相互作用はなくなり、FS315と類似したオープンな高次構造をとる(図2、3)。アクチビン・フォリスタチンは1:2のモル比率で結合する。アクチビンの二量体に2つのフォリスタチン分子が囲い込むように結合する(図2、3)。FSD1とFSD2でアクチビンのII型受容体への結合領域をふさぎ込む形をとる。FSNドメインは、主にI型受容体への結合領域をカバーしている [6]。さらにFSNドメインは、2分子目のフォリスタチンのFSD3と相互作用することで二量体に関与する[7](図3)。全体として、アクチビンのフォリスタチンへの親和性はアクチビン受容体よりも強い。こういった機構によって、アクチビンはフォリスタチンに完全に包み込まれ、受容体に結合できずシグナル伝達は遮断された状態になる[8][9][5]。
3番目のカルボキシル側のFSD3を欠損させてもアクチビン結合は保たれているが、1:1の結合になる [7]。FSNドメインを保持しつつFSD1を連結させた人為的変異体は、アクチビンとの結合は欠くが、マイオスタチンとの結合と阻害活性は保たれており筋肉量を増加させる作用を持つ [10][11][7]。
サブファミリー
TGF-βファミリーに結合するフォリスタチンのファミリー分子としては、フォリスタチン関連タンパク質3 (follistatin-like 3; FSTL3またはfollistatin related gene; FLRG)が知られている。FSTL3は263個のアミノ酸からなるペプチドホルモンで、FSTL3はフォリスタチンと異なり、N末端ドメイン(FSN)と2つのFSドメイン (FSD1、FSD2)しか待たない(図1)。TGF-βファミリーとの結合特性はフォリスタチンと類似している [12][13][14]。FSTL3の場合は、FSNでI型受容体結合部位をふさぎ、FSD1、FSD2でII型受容体との結合を阻害する。なお、FSTL3のFSD2のみでアクチビンに結合でき、アクチビンを精製することが可能である [12][15]。FSTN3のFSNはマイオスタチンとの結合に関与しリガンドの特異性に寄与する[16]。
質量分析解析によりFSTN3は血液中でマイオスタチンと結合することが報告されている [17]。アクチビンやマイオスタチンがフォリスタチン、FSTN3から遊離する機構は不明点が多いがプロテアーゼであるトロイド(Tolloid)の関与が報告されている [18]。フォリスタチンとFSTL3以外にもFSドメインを持つ分子は存在するがアクチビン結合活性は詳しく検証されていない。
立体構造の解析も進んでおり、PDBデータベースで確認することができる(表)。
| 構造 | PDB | 参考文献 |
|---|---|---|
| フォリスタチンとアクチビン複合体 | 2B0U、2ARP、2P6A | [8][19][5] |
| FSTL3(FLRG)とアクチビン複合体 | 3B4V | [20] |
| フォリスタチンとマイオスタチン複合体 | 3HH2 | [21] |
| FSTL3 (FLRG)とマイオスタチン複合体 | 3SEK | [16] |
発現
フォリスタチンのmRNAとタンパク質は、マウス、ラット、ヒトの多くの組織で発現が見られるが、卵巣、下垂体、腎臓での発現が高い。脳組織、神経系での内在性の発現は少ない。FSTN3(FLRG)のmRNAとタンパク質は、胎盤、骨髄、精巣、腎臓、骨格筋、肺での発現が高い。フォリスタチンは、アクチビン以外にGDF11を阻害する。GDF11は、嗅上皮における神経新生に関与している。アンタゴニストであるフォリスタチンを欠損させたマウスは、神経新生の劇的な減少を示す [22]。

機能
フォリスタチンは細胞外でアクチビンA、B、ABと高親和性で結合しその作用を抑制する(図4)。このことからアクチビンの作用を研究するための阻害分子として用いられることが多く有用である。上述のように、フォリスタチンは、アクチビンとモル比2:1で高親和性で結合し強く阻害する。アクチビンとの親和性と阻害活性は相関する。FS288の場合は、ヘパラン硫酸を介して細胞表層のプロテオグリカンに親和性があり、結合したアクチビンを細胞内にエンドサイトーシスの機構で取り込み分解する作用を有する[4]。FS315にはその作用はない。
フォリスタチンは、下垂体組織中での局所的なパラクリン作用でアクチビンの作用に対して阻害的に働く。アクチビンによってフォリスタチンの発現は上方制御されるが、これは一種のフィードバック機能と言える。生殖腺でもパラクライン作用でアクチビンを阻害する。なお、フォリスタチンは、マイオスタチン、GDF11に対しても阻害効果を持つ。
ノックアウトマウスの表現型
フォリスタチンの全身型のノックアウトマウスが作成されている。肺胞膨張不全のため、生後数時間で死亡するが、成長遅延、横隔膜や肋間筋量の低下、口蓋や肋骨の部分的欠損・形成異常、頬髭と歯の異常、皮膚では光沢を持ち張りのある表現型が見られる [23]。これらの表現型は、フォリスタチンがアクチビンのみならず他のTGF-βファミリーの活性を生体内で制御していることを示している。筋量低下はマイオスタチン阻害が解除されたためと推測される。
FSTL3(FLRG)のノックアウトマウスは、成体まで生存するが、膵β細胞は過形成を示し、膵島の数と大きさが増す。それに伴い、内臓脂肪量は低下し、耐糖能は改善しインスリン感受性が促進される。また、肝硬変と軽度高血圧を示すが、筋量や体重には変化は見られない [24]。
神経系とフォリスタチン
発生過程において、セグメント構造を示す後脳原基の菱形脳において、フォリスタチンは偶数の菱形脳 (r2、4、6)で発現するがr3では発現しない。このことから、Krox-20により転写抑制されると考えられている [25]。
フォリスタチンの成体の神経系での発現は低い。前脳特異的な発現を示すαCaMKIIプロモーターを用いることで作成されたフォリスタチンの過剰発現遺伝子改変マウスでは、オープンフィールドテストや明暗試験、新規物体認識試験の結果から活動性の低下と不安行動の増加が報告されている。さらに、神経新生や生存の低下が見られる。これらの表現型はアクチビン阻害によるものと考えられている [26]。アクチビンは海馬歯状回やCA1での後期長期増強(L-LTP)に必須であるが、フォリスタチンはそれを阻害する [27]。記憶は初期は海馬依存性であり、やがて皮質に移行する、記憶固定化という段階を経るが、海馬での神経新生が低下した前脳特異的過剰発現マウスでは、このプロセスが遅延し、海馬依存性の期間が長くなる[28]。
フォリスタチンやFSTN3はマイオスタチン阻害作用も強力であり、筋肥大効果が期待できる。そのため、遺伝性筋疾患の治療にこれらを利活用する研究が推進されている。フォリスタチンの骨格筋特異的な遺伝子強制発現マウスは顕著な筋肥大を示す [29]。以前より、マイオスタチンノックアウトマウスは顕著な筋肥大を示すことが知られていたが、フォリスタチン過剰発現マウスと組合わせることでさらに筋量が増す [30][31][32]。これらの結果から、フォリスタチンは生体内でマイオスタチンとアクチビンの両者を阻害し、アクチビンも筋量調節に寄与すると考察されている。血液中のアクチビン濃度は齧歯類より霊長類の方が4倍程度高いが共に1ng/ml以下である。逆に、マイオスタチンの血液濃度はマウスでは40ng/ml程度と高いが霊長類やラットでは10ng/ml以下とされている。マウスでマイオスタチン阻害が強力に筋肥大を示すのはその高い血中濃度のためと推測されている [33]。様々なアプローチによるマイオスタチン阻害が考えられるが、フォリスタチンやFSTN3もその有力な候補である[34][35][36][37]。
ヒトへのマイオスタチン阻害剤投与の治験では筋肥大効果は奏功を示していないのが現状である [38][39]。ヒトを含めた霊長類では、筋肉量の調節はアクチビンとマイオスタチンの両者によって制御されている可能性が示唆されており、アクチビンAがより重要ではないかと考察されている [33]。
疾患との関わり
フォリスタチンが持つマイオスタチンおよびアクチビン阻害活性を利用して、筋ジストロフィーなどの神経筋疾患への治療応用が期待されている。ウイルスベクターを用いた方法や安定性を増すために免疫グロブリンとの融合タンパク質や一部のドメイン構造のみを利用した手法が試みられている [40][41]。脊髄性筋萎縮症 (spinal muscular atrophy; SMA)やサルコペニアに対してマイオスタチン/アクチビン阻害療法が有効である可能性があり [42][43][44]、今後の研究の展開に期待したい。
関連語
参考文献
- ↑
Ueno, N., Ling, N., Ying, S.Y., Esch, F., Shimasaki, S., & Guillemin, R. (1987).
Isolation and partial characterization of follistatin: a single-chain Mr 35,000 monomeric protein that inhibits the release of follicle-stimulating hormone. Proceedings of the National Academy of Sciences of the United States of America, 84(23), 8282-6. [PubMed:3120188] [PMC] [WorldCat] [DOI] - ↑
Nakamura, T., Takio, K., Eto, Y., Shibai, H., Titani, K., & Sugino, H. (1990).
Activin-binding protein from rat ovary is follistatin. Science (New York, N.Y.), 247(4944), 836-8. [PubMed:2106159] [WorldCat] [DOI] - ↑
Shimasaki, S., Koga, M., Esch, F., Cooksey, K., Mercado, M., Koba, A., ..., & Guillemin, R. (1988).
Primary structure of the human follistatin precursor and its genomic organization. Proceedings of the National Academy of Sciences of the United States of America, 85(12), 4218-22. [PubMed:3380788] [PMC] [WorldCat] [DOI] - ↑ 4.0 4.1
Hashimoto, O., Nakamura, T., Shoji, H., Shimasaki, S., Hayashi, Y., & Sugino, H. (1997).
A novel role of follistatin, an activin-binding protein, in the inhibition of activin action in rat pituitary cells. Endocytotic degradation of activin and its acceleration by follistatin associated with cell-surface heparan sulfate. The Journal of biological chemistry, 272(21), 13835-42. [PubMed:9153241] [WorldCat] [DOI] - ↑ 5.0 5.1 5.2
Lerch, T.F., Shimasaki, S., Woodruff, T.K., & Jardetzky, T.S. (2007).
Structural and biophysical coupling of heparin and activin binding to follistatin isoform functions. The Journal of biological chemistry, 282(21), 15930-9. [PubMed:17409095] [WorldCat] [DOI] - ↑
Greenwald, J., Vega, M.E., Allendorph, G.P., Fischer, W.H., Vale, W., & Choe, S. (2004).
A flexible activin explains the membrane-dependent cooperative assembly of TGF-beta family receptors. Molecular cell, 15(3), 485-9. [PubMed:15304227] [WorldCat] [DOI] - ↑ 7.0 7.1 7.2
Cash, J.N., Angerman, E.B., Keutmann, H.T., & Thompson, T.B. (2012).
Characterization of follistatin-type domains and their contribution to myostatin and activin A antagonism. Molecular endocrinology (Baltimore, Md.), 26(7), 1167-78. [PubMed:22593183] [PMC] [WorldCat] [DOI] - ↑ 8.0 8.1
Thompson, T.B., Lerch, T.F., Cook, R.W., Woodruff, T.K., & Jardetzky, T.S. (2005).
The structure of the follistatin:activin complex reveals antagonism of both type I and type II receptor binding. Developmental cell, 9(4), 535-43. [PubMed:16198295] [WorldCat] [DOI] - ↑
Harrington, A.E., Morris-Triggs, S.A., Ruotolo, B.T., Robinson, C.V., Ohnuma, S., & Hyvönen, M. (2006).
Structural basis for the inhibition of activin signalling by follistatin. The EMBO journal, 25(5), 1035-45. [PubMed:16482217] [PMC] [WorldCat] [DOI] - ↑
Nakatani, M., Kokubo, M., Ohsawa, Y., Sunada, Y., & Tsuchida, K. (2011).
Follistatin-derived peptide expression in muscle decreases adipose tissue mass and prevents hepatic steatosis. American journal of physiology. Endocrinology and metabolism, 300(3), E543-53. [PubMed:21205933] [WorldCat] [DOI] - ↑
Nakatani, M., Takehara, Y., Sugino, H., Matsumoto, M., Hashimoto, O., Hasegawa, Y., ..., & Tsuchida, K. (2008).
Transgenic expression of a myostatin inhibitor derived from follistatin increases skeletal muscle mass and ameliorates dystrophic pathology in mdx mice. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 22(2), 477-87. [PubMed:17893249] [WorldCat] [DOI] - ↑ 12.0 12.1
Tsuchida, K., Arai, K.Y., Kuramoto, Y., Yamakawa, N., Hasegawa, Y., & Sugino, H. (2000).
Identification and characterization of a novel follistatin-like protein as a binding protein for the TGF-beta family. The Journal of biological chemistry, 275(52), 40788-96. [PubMed:11010968] [WorldCat] [DOI] - ↑
Sidis, Y., Mukherjee, A., Keutmann, H., Delbaere, A., Sadatsuki, M., & Schneyer, A. (2006).
Biological activity of follistatin isoforms and follistatin-like-3 is dependent on differential cell surface binding and specificity for activin, myostatin, and bone morphogenetic proteins. Endocrinology, 147(7), 3586-97. [PubMed:16627583] [WorldCat] [DOI] - ↑
Tsuchida, K., Matsuzaki, T., Yamakawa, N., Liu, Z., & Sugino, H. (2001).
Intracellular and extracellular control of activin function by novel regulatory molecules. Molecular and cellular endocrinology, 180(1-2), 25-31. [PubMed:11451568] [WorldCat] [DOI] - ↑
Arai, K.Y., Tsuchida, K., Li, C., Watanabe, G., Sugino, H., Taya, K., & Nishiyama, T. (2006).
Purification of recombinant activin A using the second follistatin domain of follistatin-related gene (FLRG). Protein expression and purification, 49(1), 78-82. [PubMed:16737827] [WorldCat] [DOI] - ↑ 16.0 16.1
Cash, J.N., Angerman, E.B., Kattamuri, C., Nolan, K., Zhao, H., Sidis, Y., ..., & Thompson, T.B. (2012).
Structure of myostatin·follistatin-like 3: N-terminal domains of follistatin-type molecules exhibit alternate modes of binding. The Journal of biological chemistry, 287(2), 1043-53. [PubMed:22052913] [PMC] [WorldCat] [DOI] - ↑
Hill, J.J., Davies, M.V., Pearson, A.A., Wang, J.H., Hewick, R.M., Wolfman, N.M., & Qiu, Y. (2002).
The myostatin propeptide and the follistatin-related gene are inhibitory binding proteins of myostatin in normal serum. The Journal of biological chemistry, 277(43), 40735-41. [PubMed:12194980] [WorldCat] [DOI] - ↑
Walker, R.G., McCoy, J.C., Czepnik, M., Mills, M.J., Hagg, A., Walton, K.L., ..., & Thompson, T.B. (2018).
Molecular characterization of latent GDF8 reveals mechanisms of activation. Proceedings of the National Academy of Sciences of the United States of America, 115(5), E866-E875. [PubMed:29348202] [PMC] [WorldCat] [DOI] - ↑
Lin, S.J., Lerch, T.F., Cook, R.W., Jardetzky, T.S., & Woodruff, T.K. (2006).
The structural basis of TGF-beta, bone morphogenetic protein, and activin ligand binding. Reproduction (Cambridge, England), 132(2), 179-90. [PubMed:16885528] [WorldCat] [DOI] - ↑
Stamler, R., Keutmann, H.T., Sidis, Y., Kattamuri, C., Schneyer, A., & Thompson, T.B. (2008).
The structure of FSTL3.activin A complex. Differential binding of N-terminal domains influences follistatin-type antagonist specificity. The Journal of biological chemistry, 283(47), 32831-8. [PubMed:18768470] [PMC] [WorldCat] [DOI] - ↑
Cash, J.N., Rejon, C.A., McPherron, A.C., Bernard, D.J., & Thompson, T.B. (2009).
The structure of myostatin:follistatin 288: insights into receptor utilization and heparin binding. The EMBO journal, 28(17), 2662-76. [PubMed:19644449] [PMC] [WorldCat] [DOI] - ↑
Wu, H.H., Ivkovic, S., Murray, R.C., Jaramillo, S., Lyons, K.M., Johnson, J.E., & Calof, A.L. (2003).
Autoregulation of neurogenesis by GDF11. Neuron, 37(2), 197-207. [PubMed:12546816] [WorldCat] [DOI] - ↑
Matzuk, M.M., Lu, N., Vogel, H., Sellheyer, K., Roop, D.R., & Bradley, A. (1995).
Multiple defects and perinatal death in mice deficient in follistatin. Nature, 374(6520), 360-3. [PubMed:7885475] [WorldCat] [DOI] - ↑
Mukherjee, A., Sidis, Y., Mahan, A., Raher, M.J., Xia, Y., Rosen, E.D., ..., & Schneyer, A.L. (2007).
FSTL3 deletion reveals roles for TGF-beta family ligands in glucose and fat homeostasis in adults. Proceedings of the National Academy of Sciences of the United States of America, 104(4), 1348-53. [PubMed:17229845] [PMC] [WorldCat] [DOI] - ↑
Seitanidou, T., Schneider-Maunoury, S., Desmarquet, C., Wilkinson, D.G., & Charnay, P. (1997).
Krox-20 is a key regulator of rhombomere-specific gene expression in the developing hindbrain. Mechanisms of development, 65(1-2), 31-42. [PubMed:9256343] [WorldCat] [DOI] - ↑
Ageta, H., Murayama, A., Migishima, R., Kida, S., Tsuchida, K., Yokoyama, M., & Inokuchi, K. (2008).
Activin in the brain modulates anxiety-related behavior and adult neurogenesis. PloS one, 3(4), e1869. [PubMed:18382659] [PMC] [WorldCat] [DOI] - ↑
Ageta, H., Ikegami, S., Miura, M., Masuda, M., Migishima, R., Hino, T., ..., & Inokuchi, K. (2010).
Activin plays a key role in the maintenance of long-term memory and late-LTP. Learning & memory (Cold Spring Harbor, N.Y.), 17(4), 176-85. [PubMed:20332189] [WorldCat] [DOI] - ↑
Kitamura, T., Saitoh, Y., Takashima, N., Murayama, A., Niibori, Y., Ageta, H., ..., & Inokuchi, K. (2009).
Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell, 139(4), 814-27. [PubMed:19914173] [WorldCat] [DOI] - ↑
Lee, S.J., & McPherron, A.C. (2001).
Regulation of myostatin activity and muscle growth. Proceedings of the National Academy of Sciences of the United States of America, 98(16), 9306-11. [PubMed:11459935] [PMC] [WorldCat] [DOI] - ↑
McPherron, A.C., Lawler, A.M., & Lee, S.J. (1997).
Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature, 387(6628), 83-90. [PubMed:9139826] [WorldCat] [DOI] - ↑
Lee, S.J. (2007).
Quadrupling muscle mass in mice by targeting TGF-beta signaling pathways. PloS one, 2(8), e789. [PubMed:17726519] [PMC] [WorldCat] [DOI] - ↑
Lee, S.J., Lee, Y.S., Zimmers, T.A., Soleimani, A., Matzuk, M.M., Tsuchida, K., ..., & Barton, E.R. (2010).
Regulation of muscle mass by follistatin and activins. Molecular endocrinology (Baltimore, Md.), 24(10), 1998-2008. [PubMed:20810712] [PMC] [WorldCat] [DOI] - ↑ 33.0 33.1
Latres, E., Mastaitis, J., Fury, W., Miloscio, L., Trejos, J., Pangilinan, J., ..., & Gromada, J. (2017).
Activin A more prominently regulates muscle mass in primates than does GDF8. Nature communications, 8, 15153. [PubMed:28452368] [PMC] [WorldCat] [DOI] - ↑
Tsuchida, K., Nakatani, M., Hitachi, K., Uezumi, A., Sunada, Y., Ageta, H., & Inokuchi, K. (2009).
Activin signaling as an emerging target for therapeutic interventions. Cell communication and signaling : CCS, 7, 15. [PubMed:19538713] [PMC] [WorldCat] [DOI] - ↑
Saitoh, M., Takayama, K., Hitachi, K., Taguchi, A., Taniguchi, A., Tsuchida, K., & Hayashi, Y. (2020).
Discovery of a follistatin-derived myostatin inhibitory peptide. Bioorganic & medicinal chemistry letters, 30(3), 126892. [PubMed:31874826] [WorldCat] [DOI] - ↑
Lee, S.J. (2021).
Targeting the myostatin signaling pathway to treat muscle loss and metabolic dysfunction. The Journal of clinical investigation, 131(9). [PubMed:33938454] [PMC] [WorldCat] [DOI] - ↑
Ozawa, T., Morikawa, M., Morishita, Y., Ogikubo, K., Itoh, F., Koinuma, D., ..., & Miyazono, K. (2021).
Systemic administration of monovalent follistatin-like 3-Fc-fusion protein increases muscle mass in mice. iScience, 24(5), 102488. [PubMed:34113826] [PMC] [WorldCat] [DOI] - ↑
Wagner, K.R. (2020).
The elusive promise of myostatin inhibition for muscular dystrophy. Current opinion in neurology, 33(5), 621-628. [PubMed:32773450] [WorldCat] [DOI] - ↑
Suh, J., & Lee, Y.S. (2020).
Myostatin Inhibitors: Panacea or Predicament for Musculoskeletal Disorders? Journal of bone metabolism, 27(3), 151-165. [PubMed:32911580] [PMC] [WorldCat] [DOI] - ↑
Kota, J., Handy, C.R., Haidet, A.M., Montgomery, C.L., Eagle, A., Rodino-Klapac, L.R., ..., & Kaspar, B.K. (2009).
Follistatin gene delivery enhances muscle growth and strength in nonhuman primates. Science translational medicine, 1(6), 6ra15. [PubMed:20368179] [PMC] [WorldCat] [DOI] - ↑
Rodino-Klapac, L.R., Haidet, A.M., Kota, J., Handy, C., Kaspar, B.K., & Mendell, J.R. (2009).
Inhibition of myostatin with emphasis on follistatin as a therapy for muscle disease. Muscle & nerve, 39(3), 283-96. [PubMed:19208403] [PMC] [WorldCat] [DOI] - ↑
Crawford, T.O., Darras, B.T., Day, J.W., Dunaway Young, S., Duong, T., Nelson, L.L., ..., & Chyung, Y. (2024).
Safety and Efficacy of Apitegromab in Patients With Spinal Muscular Atrophy Types 2 and 3: The Phase 2 TOPAZ Study. Neurology, 102(5), e209151. [PubMed:38330285] [PMC] [WorldCat] [DOI] - ↑
Servais, L., Lair, L.L., Connolly, A.M., Byrne, B.J., Chen, K.S., Coric, V., ..., & Bechtold, C. (2024).
Taldefgrobep Alfa and the Phase 3 RESILIENT Trial in Spinal Muscular Atrophy. International journal of molecular sciences, 25(19). [PubMed:39408601] [PMC] [WorldCat] [DOI] - ↑
Bromer, F.D., Lodberg, A., Eijken, M., Andersen, C.B.F., Poulsen, M.F., Thomsen, J.S., & Brüel, A. (2025).
The Effect of Anti-Activin Receptor Type IIA and Type IIB Antibody on Muscle, Bone and Blood in Healthy and Osteosarcopenic Mice. Journal of cachexia, sarcopenia and muscle, 16(1), e13718. [PubMed:39887865] [PMC] [WorldCat] [DOI]