「リアノジン受容体」の版間の差分
細編集の要約なし |
細編集の要約なし |
(相違点なし)
| |
2021年6月14日 (月) 09:17時点における最新版
柿澤 昌
京都大学 大学院薬学研究科 生体分子認識学分野
DOI:10.14931/bsd.2541 原稿受付日:2012年9月26日 原稿完成日:2012年10月23日
担当編集委員:林 康紀(独立行政法人理化学研究所 脳科学総合研究センター)
英語名:Ryanodine receptor 英語略名:RyR
リアノジン受容体は細胞内カルシウム貯蔵部位である小胞体膜上に存在するカルシウムチャネルであり、その名は、植物アルカロイドであるリアノジンが結合することに由来する。小胞体からのカルシウム放出を担うことから、同じく小胞体膜上に存在するカルシウムチャネルであるイノシトール1,4,5-三リン酸受容体とともに、カルシウム放出チャネルとも呼ばれ、細胞内カルシウム濃度調節に関与する。三種類のサブタイプが存在し、それぞれ異なった分布を示すが、脳においては三種類全ての発現が見られる。三種類とも細胞質のカルシウム濃度上昇より活性化されるが、1型リアノジン受容体はS-ニトロシル化などの化学修飾や脱分極(骨格筋)によっても活性化される。また三種類のサブタイプ全てに対して遺伝子欠損マウスが作成されているが、1型RyR欠損マウスは出生致死、2型リアノジン受容体欠損マウスは胎生致死を示す。3型リアノジン受容体欠損マウスのみ生後も生存・成熟するため解析が可能であり、脳機能への関与についての報告が存在する。他にも主に薬理学的なアプローチにより、リアノジン受容体のシナプス可塑性・神経細胞興奮性などの生理学的役割及び興奮性神経細胞死などの病態生理学的役割が示されている。


| Ryanodine receptor 2 (cardiac) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||
| Symbols | RYR2; ARVC2; ARVD2; RYR-2; RyR; VTSIP | ||||||||||||
| External IDs | OMIM: 180902 MGI: 99685 HomoloGene: 37423 IUPHAR: RyR2 GeneCards: RYR2 Gene | ||||||||||||
| |||||||||||||
| Orthologs | |||||||||||||
| Species | Human | Mouse | |||||||||||
| Entrez | 6262 | 20191 | |||||||||||
| Ensembl | ENSG00000198626 | ENSMUSG00000021313 | |||||||||||
| UniProt | Q92736 | E9Q401 | |||||||||||
| RefSeq (mRNA) | NM_001035.2 | NM_023868.2 | |||||||||||
| RefSeq (protein) | NP_001026.2 | NP_076357.2 | |||||||||||
| Location (UCSC) |
Chr 1: 237.21 – 238.13 Mb |
Chr 13: 11.55 – 12.11 Mb | |||||||||||
| PubMed search | [3] | [4] | |||||||||||
| Ryanodine receptor 3 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||
| Identifiers | |||||||||||||
| Symbol | RYR3 | ||||||||||||
| External IDs | OMIM: 180903 MGI: 99684 HomoloGene: 68151 IUPHAR: RyR3 GeneCards: RYR3 Gene | ||||||||||||
| |||||||||||||
| Orthologs | |||||||||||||
| Species | Human | Mouse | |||||||||||
| Entrez | 6263 | 20192 | |||||||||||
| Ensembl | ENSG00000198838 | ENSMUSG00000057378 | |||||||||||
| UniProt | Q15413 | A2AGL3 | |||||||||||
| RefSeq (mRNA) | NM_001036.3 | NM_177652.2 | |||||||||||
| RefSeq (protein) | NP_001027.3 | NP_808320.2 | |||||||||||
| Location (UCSC) |
Chr 15: 33.6 – 34.16 Mb |
Chr 2: 112.63 – 113.22 Mb | |||||||||||
| PubMed search | [5] | [6] | |||||||||||
歴史
カルシウムイオン(Ca2+)は普遍的かつ基本的なシグナル伝達を担うセカンドメッセンジャーであり、極めて多くの生命現象に関与する。細胞内におけるカルシウムシグナル形成は、細胞膜に存在するカルシウムチャネルを介して細胞外から細胞内へのカルシウムの流入によるものと、細胞内カルシウムストア(小胞体)からカルシウム放出チャネルを介して細胞質へ放出される2通りの経路による(図1)。カルシウム誘発性カルシウム放出(Ca2+-induced Ca2+ release; CICR)は、細胞質側のカルシウム濃度上昇が細胞内ストアから細胞質へのカルシウム放出を促進する現象であり、骨格筋で最初に見出された[1]。その後、同様の現象が多くの興奮性細胞において見られたことから、CICRは細胞内カルシウムシグナルを増幅するための普遍的な機構であると考えられるようになり、CICRの分子実体であるCICRチャネルの薬理学的性質が調べられた。その結果、植物アルカロイドであるリアノジンがCICRチャネルに特異的に結合し、低濃度ではチャネルを開口状態に固定する薬物であることが示された。
引き続き、標識リアノジンを用いた結合活性を指標に、骨格筋よりCICRチャネル、即ちリアノジン受容体(RyR)が精製された[2]。その後の遺伝子クローニングにより、少なくとも硬骨魚類以上の脊椎動物では、別々の遺伝子にコードされる3種類のRyRサブタイプが存在することが判明し、それぞれ、1型/骨格筋型(RyR1)、2型/心筋型(RyR2)、3型/脳型(RyR3)と呼ばれる[3][4]。各サブタイプは互いに65%程度のアミノ酸配列相同性を示すが、異なる組織分布・脳内分布を示す[5][6]。一方、線虫[7]、ショウジョウバエ[8]においては、どのタイプにも属さないRyR相同物が同定されており、無脊椎動物においては単一遺伝子にコードされていたものが、脊椎動物において組織分布や機能的役割が異なる3種のサブタイプに分子進化したと推測されている。
-
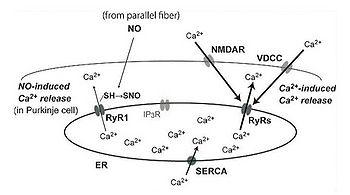 図1.リアノジン受容体を介するシグナル系
図1.リアノジン受容体を介するシグナル系
脳の神経細胞におけるリアノジン受容体(RyRs)を介するシグナル伝達。海馬の錐体細胞ではNMDA型グルタミン酸受容体(NMDAR)、小脳のプルキンエ細胞では電位依存症カルシウムチャネル(VDCC)を介する細胞外からのカルシウム流入による細胞内カルシウム濃度上昇により、Ca2+-induced Ca2+ releaseが起こる。一方、小脳プルキンエ細胞では一酸化窒素(NO)による1型RyR(RyR1)のS-ニトロシル化によりNO-induced Ca2+releaseも起こる。ER:endoplasmic reticulum; IP3R:inositol 1,4,5 tris phosphate receptor; SERCA: sarco/endoplasmic reticulum Ca2+ ATPase.

分子構造
RyRは約5,000アミノ酸残基より構成される分子量約550kDaの巨大タンパク質であり、そのホモ4量体により機能的なカルシウム放出チャネルが形成される。RyR分子内では、アミノ末端側約4,500アミノ酸が細胞質側に大きく張り出した、いわゆるfoot構造に対応する領域を形成し、4本の推定膜貫通セグメントを含む残りのカルボキシル末端がチャネル領域を形成する(図2)。同じくカルシウム放出チャネルであるIP3受容体とRyRは、カルボキシル末端の膜貫通セグメントを含む約200アミノ酸において特に高い配列相同性を示し、この部分がカルシウム放出チャネルとして共通するイオン透過性に関連した機能に関与していると推測される。実際に、RyR1のカルボキシル末側の約1,000アミノ酸残基よりなる領域のみでリアノジン感受性を有する基本的なカルシウムチャネルが形成されることが、脂質二重膜を用いた再構築実験により示されている。この再構築系においてC末端領域が形成するチャネルは、陽イオン選択性、細胞質側カルシウムによる活性化機構、リアノジン感受性を保持する一方で、全長RyR1チャネルに見られる高濃度のカルシウムによる不活性化機構を欠く。また、RyRサブタイプ間での一次構造の比較により、D1、D2、D3と呼ばれる相同性が顕著に低い領域が見られる。RyR1のcDNA発現実験系を用いた研究により、CICRによるチャネル活性化のカルシウム結合部位はD1領域、チャネルを不活性化する高濃度カルシウムの結合部位はD3領域に存在するとされている[9][10](図3)。
-
 図2.リアノジン受容体の三次元構造
図2.リアノジン受容体の三次元構造 -
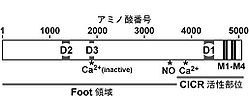 図3.リアノジン受容体の一次構造
図3.リアノジン受容体の一次構造


各サブタイプの体内分布
RyR1
RyR1は骨格筋において極めて高レベルの発現が見られる。また、小脳プルキンエ細胞を始めとする脳内各部位でも発現が見られる他、食道、精巣でも他の部位に比較して高レベルの発現が見られる。小脳プルキンエ細胞においては、高レベルの発現が見られるとともに3つのサブタイプの中でも最も発現量が多い。海馬歯状回でも高レベルの発現が見られるが、歯状回においてはRyR2の発現量が最も多い。さらに、線条体、嗅球僧帽細胞などでも、他の領域と比較して高レベルの発現が見られる。
RyR2
RyR2は心筋細胞で極めて高レベルの発現が見られる他、平滑筋・肺・脳でも高レベルの発現が見られる。脳内の様々な部位において広く発現が見られるが、特に海馬(歯状回・CA領域)、大脳皮質、小脳顆粒細胞層、嗅球顆粒細胞層などで高レベルの発現が見られる。
RyR3
RyR3は脳cDNAライブラリーからのクローニングにより存在が明らかになったが、脳以外にも平滑筋、骨格筋、一部の上皮細胞やリンパ球培養細胞などにおいて低レベルの発現が見られる。海馬CA領域で高レベルの発現が見られる。海馬歯状回、線条体などでも比較的発現レベルが高い。
尚、各サブタイプの体内分布に関する詳細については、Giannini et al. (1995) [6] を参考にされたい。
活性調節因子
RyRの活性調節因子に関しては、主に筋細胞や再構築系を用いた多数の研究報告がある。詳細については、これらの報告をまとめた総説を参考にされたい[9][10][11]。
カルシウムイオン
細胞質に存在するカルシウムイオンは、濃度依存的に全てのRyRサブタイプに共通して作用する調節因子である。サブμMからμMの範囲における細胞質側カルシウムはRyRを開口させ細胞内カルシウムストアである小胞体からカルシウムを遊離させる、いわゆるカルシウム依存性(誘発性)カルシウム放出(CICR)現象をお引き起こす。一方、より高濃度のmMレベルのカルシウムはチャネル活性を抑制する。また、小胞体内腔側のカルシウムによる活性化も見られる。心筋では、脱分極によるL型カルシウムチャネル(Cv1.2)の開口により細胞外からカルシウムが流入し、RyR2を開口させカルシウム放出を引き起こし、細胞内カルシウムシグナルを増幅するが、中枢神経系においても、小脳プルキンエ細胞では電位依存性のP/Q型カルシウムチャネル、海馬錐体細胞ではNMDA型グルタミン酸受容体を介するカルシウム流入によりRyRが活性化されカルシウム放出が誘導されることが示唆されている。
脱分極
骨格筋においてはL型カルシウムチャネル(Cav1.1)とRyR1が機械的にカップリングし、興奮収縮連関において細胞膜が脱分極するとL型カルシウムチャネルのコンフォーメーションが変化し、タンパク質-タンパク質相互作用を介してRyR1が活性化されると考えられている。一方、中枢神経系における機械的なカップリングに関しては、多くの点が不明である。
生理活性物質
RyRの内在性の因子による活性調節として、カルモジュリンによる濃度依存的な活性化と不活性化、ATPを含むアデニンヌクレオチド、プロテインキナーゼAなどのリン酸化酵素、cADPリボースなどによる活性化、および免疫抑制薬であるFK506の結合タンパク質FKBP12によるRyR2チャネルの静止および開口状態を安定化する作用が示唆されている。さらに一酸化窒素(NO)によるS-ニトロシル化がRyR1の開口確率を上昇させることがin vitroの実験系により示されていたが、つい最近、小脳プルキンエ細胞において内因性NOの作用によりRyR1を介したカルシウム放出が誘導されることが示された(一酸化窒素依存的カルシウム放出の項 参照)。
薬物
リアノジンは低濃度では開状態のRyRに結合し、サブコンダクタンス状態に開口固定しカルシウム遊離を引き起こすが、高濃度ではRyR活性を抑制する。また、カフェインによるRyRの活性化、ダントロレン dantrolene による抑制(主にRyR1、RyR3に対する作用)が知られている。
機能
薬理学的な刺激により、脳における機能的RyRの存在、あるいはRyRを介するカルシウム放出の存在を示した報告は、現在では多数存在する。しかし、生理的な刺激によるRyRの活性化を示した報告は、薬理学的刺激による報告に比べると遥かに少なく、さらに、シナプス可塑性や個体の行動の様な機能的役割と関連付けたものは限られたものになり、RyRの脳における機能的役割は、未だ解明の途上にあると言える[11]。
ノックアウトマウスの表現型
RyRの機能的役割解明を困難なものにしている主要な原因として、遺伝子欠損動物の致死性が挙げられる。上述の通り、哺乳類の脳の多くの領域では、複数のRyRサブタイプが重複して発現しているが、RyR1欠損マウス、RyR2欠損マウスはそれぞれ単独で、出生致死、胎生致死を示す。したがって、複数のRyRサブタイプ遺伝子の二重もしくは三重欠損マウスが成熟しないことは自明であり、全身レベルでの遺伝子欠損マウスを用いたアプローチによっては、脳におけるRyRの機能は困難である。しかし、RyR3欠損マウスは生後も生存・成熟するため、その解析結果の報告が存在する[12]。
RyR1欠損マウス
RyR2欠損マウス
- RyR2欠損マウスは、心拍動の開始直後の胎生10日ごろに心筋細胞の小胞体カルシウム過剰負荷により心不全となり死亡する[14]。
RyR3欠損マウス
- RyR3欠損マウスはほぼ正常に発育し重篤な異常は認められないが、これまでに自発的運動活性の亢進、社会的接触行動の減少、恐怖条件付け反応の低下が報告され、その神経系での重要性が示唆されている。また、海馬CA1領域において、穏やかな刺激で誘導されたLTPの維持が阻害されるとの報告がある一方で、同じく海馬領域におけるLTPの誘導閾値が低下するとの報告もある。ただし、RyR3欠損マウスの軽度な中枢機能異常に関しては、重複して発現する他のサブタイプによる補完作用を考慮する必要がある[15][16]。
シナプス前終末における機能
海馬CA3領域の苔状線維軸索(シナプス前終末よりも軸索起始部寄りの部分)においては、電位依存性カルシウムチャネルによるカルシウムシグナルがRyR1によるCICR機構を介して増幅されることにより、高頻度刺激に神経伝達物資の放出が増強されることが示されており、シナプス前終末における可塑性へのRyRの関与も示唆されている[17]。
一酸化窒素依存的カルシウム放出
脂質二重膜に発現させたRyR1の開口確率がNOの作用により上昇することは以前より知られていたが、この現象が生細胞で内因性のNOの作用により起こること、およびその機能的意義については長いこと不明であった。しかし、小脳平行線維-プルキンエ細胞シナプスにおけるNO依存的LTPがプルキンエ細胞内の細胞内カルシウムシグナルにも依存的であることが判明したことから、プルキンエ細胞内でのNOとカルシウムとの関連性について解明が進み、神経活動によって産生放出された内因性のNOがRyR1を活性化することでカルシウム放出が誘導される現象、NO依存的カルシウム放出(NO-induced Ca2+ release; NICR)が発見された。このNICRはウサギRyR1における3635位のシステイン(マウスでは3636位に相当)がNOによりS-ニトロシル化されることで誘導されると推測されている。また、NO合成酵素の発現は平行線維では見られるがプルキンエ細胞では見られないことから、平行線維活動により産生放出されたNOがプルキンエ細胞内のRyR1を活性化すると考えられている。これまでに、NICRの小脳平行線維-プルキンエ細胞シナプスにおけるLTPへの関与、および、中大脳動脈の虚血再灌流による大脳皮質の神経細胞死への関与が示唆されている[18]。
脳型ジャンクトフィリン欠損マウスの解析
骨格筋興奮収縮連関における電位依存性カルシウムチャネル(L型チャネル)とRyRとの機能共役のためには、細胞膜と小胞体膜とが近接した結合膜構造の形成が必要であるが、この結合膜構造に関与する分子として単離同定されたジャンクトフィリンの遺伝子欠損マウスからも、脳におけるRyRの機能的役割が示唆されている[19]。詳細はジャンクトフィリンの項目参照。
疾患との関連
悪性高熱症
Malignant hyperthermia; MH
骨格筋においてRyR1は上述のように脱分極によるCav1.1の活性化をトリガーとして活性化するが、特殊条件下ではCICR機構によるチャネル開口も起こし得る。MHはハロタンなどの吸入全身麻酔により発生する全身性の筋硬縮と発熱を特徴とする、RyR1によるCICR機構が著しく亢進した病態である。MHの原因となる変異型RyR1は、通常よりも低い細胞質カルシウム濃度においてCICR機構によるチャネルの活性化が生じる。したがって、RyRへの刺激作用を有する吸入麻酔薬に敏感に反応することで小胞体カルシウム放出が誘導され、その結果、全身の筋組織が硬縮し熱を産生する。MHのアミノ酸点変異は、RyR1のCICRチャネルとしてのカルシウム感受性をより高める変異であり、世界各地で発症した悪性高熱症についての様々な点変異が報告されている[9][10]。
カテコールアミン誘発性多形性心室頻拍
Catecholaminergic polymorphic ventricular tachycardia; CPVT
本症候群は精神的または身体的ストレス時に交感神経活動が亢進することにより誘発される多形性心室頻拍の反復出現を特徴とする疾患である。このCPVP家系には急死例が多く、常染色体性優性遺伝形式をとり、その原因遺伝子として心臓RyR2の変異が関与する。交感神経によるβ受容体刺激の心臓収縮の増強作用の機序の1つとして、心筋細胞内でcAMP依存性リン酸化酵素によるRyR2のリン酸化・活性化を介して小胞体カルシウム放出を亢進することが知られている。悪性高熱症と同様に、CPVP家系で見出される遺伝子変異はRyR2をより活性型に誘導する変異であると考えられ、β受容体刺激時に過剰にRyR2が活性化することが致死的な頻拍を誘導することが示唆されている。欧米各国での発症例についてRyR2上での様々な点変異が報告されている[9][10]。
タンパク質機能との関わり
RyR1とRyR2の一次構造上にMHとCPVPにて報告された遺伝子変異をマップすると、アミノ末端部位、中央部位およびカルボキシル末端部位に集中することが判る。基本的なCICRチャネル機能を内蔵するカルボキシル末端部位に加えて、両症候群の変異が集約されるアミノ末端部位と中央部位もRyRチャネルの開閉制御に極めて重要な機能を有することが示されている。
関連項目
参考文献
- ↑
Endo, M., Tanaka, M., & Ogawa, Y. (1970).
Calcium induced release of calcium from the sarcoplasmic reticulum of skinned skeletal muscle fibres. Nature, 228(5266), 34-6. [PubMed:5456208] [WorldCat] [DOI] - ↑
Lai, F.A., Erickson, H.P., Rousseau, E., Liu, Q.Y., & Meissner, G. (1988).
Purification and reconstitution of the calcium release channel from skeletal muscle. Nature, 331(6154), 315-9. [PubMed:2448641] [WorldCat] [DOI] - ↑
Takeshima, H. (1993).
Primary structure and expression from cDNAs of the ryanodine receptor. Annals of the New York Academy of Sciences, 707, 165-77. [PubMed:9137551] [WorldCat] [DOI] - ↑
Takeshima, H. (2003).
Ryanodine receptor and junctional membrane structure. Nihon yakurigaku zasshi. Folia pharmacologica Japonica, 121(4), 203-10. [PubMed:12777839] [WorldCat] [DOI] - ↑
Hakamata, Y., Nakai, J., Takeshima, H., & Imoto, K. (1992).
Primary structure and distribution of a novel ryanodine receptor/calcium release channel from rabbit brain. FEBS letters, 312(2-3), 229-35. [PubMed:1330694] [WorldCat] [DOI] - ↑ 6.0 6.1
Giannini, G., Conti, A., Mammarella, S., Scrobogna, M., & Sorrentino, V. (1995).
The ryanodine receptor/calcium channel genes are widely and differentially expressed in murine brain and peripheral tissues. The Journal of cell biology, 128(5), 893-904. [PubMed:7876312] [PMC] [WorldCat] [DOI] - ↑
Sakube, Y., Ando, H., & Kagawa, H. (1997).
An abnormal ketamine response in mutants defective in the ryanodine receptor gene ryr-1 (unc-68) of Caenorhabditis elegans. Journal of molecular biology, 267(4), 849-64. [PubMed:9135117] [WorldCat] [DOI] - ↑
Takeshima, H., Nishi, M., Iwabe, N., Miyata, T., Hosoya, T., Masai, I., & Hotta, Y. (1994).
Isolation and characterization of a gene for a ryanodine receptor/calcium release channel in Drosophila melanogaster. FEBS letters, 337(1), 81-7. [PubMed:8276118] [WorldCat] [DOI] - ↑ 9.0 9.1 9.2 9.3
Zalk, R., Lehnart, S.E., & Marks, A.R. (2007).
Modulation of the ryanodine receptor and intracellular calcium. Annual review of biochemistry, 76, 367-85. [PubMed:17506640] [WorldCat] [DOI] - ↑ 10.0 10.1 10.2 10.3
Lanner, J.T., Georgiou, D.K., Joshi, A.D., & Hamilton, S.L. (2010).
Ryanodine receptors: structure, expression, molecular details, and function in calcium release. Cold Spring Harbor perspectives in biology, 2(11), a003996. [PubMed:20961976] [PMC] [WorldCat] [DOI] - ↑ 11.0 11.1
Verkhratsky, A. (2005).
Physiology and pathophysiology of the calcium store in the endoplasmic reticulum of neurons. Physiological reviews, 85(1), 201-79. [PubMed:15618481] [WorldCat] [DOI] - ↑
Kushnir, A., Betzenhauser, M.J., & Marks, A.R. (2010).
Ryanodine receptor studies using genetically engineered mice. FEBS letters, 584(10), 1956-65. [PubMed:20214899] [PMC] [WorldCat] [DOI] - ↑
Takeshima, H., Iino, M., Takekura, H., Nishi, M., Kuno, J., Minowa, O., ..., & Noda, T. (1994).
Excitation-contraction uncoupling and muscular degeneration in mice lacking functional skeletal muscle ryanodine-receptor gene. Nature, 369(6481), 556-9. [PubMed:7515481] [WorldCat] [DOI] - ↑
Takeshima, H., Komazaki, S., Hirose, K., Nishi, M., Noda, T., & Iino, M. (1998).
Embryonic lethality and abnormal cardiac myocytes in mice lacking ryanodine receptor type 2. The EMBO journal, 17(12), 3309-16. [PubMed:9628868] [PMC] [WorldCat] [DOI] - ↑
Futatsugi, A., Kato, K., Ogura, H., Li, S.T., Nagata, E., Kuwajima, G., ..., & Mikoshiba, K. (1999).
Facilitation of NMDAR-independent LTP and spatial learning in mutant mice lacking ryanodine receptor type 3. Neuron, 24(3), 701-13. [PubMed:10595520] [WorldCat] [DOI] - ↑
Shimuta, M., Yoshikawa, M., Fukaya, M., Watanabe, M., Takeshima, H., & Manabe, T. (2001).
Postsynaptic modulation of AMPA receptor-mediated synaptic responses and LTP by the type 3 ryanodine receptor. Molecular and cellular neurosciences, 17(5), 921-30. [PubMed:11358488] [WorldCat] [DOI] - ↑
Shimizu, H., Fukaya, M., Yamasaki, M., Watanabe, M., Manabe, T., & Kamiya, H. (2008).
Use-dependent amplification of presynaptic Ca2+ signaling by axonal ryanodine receptors at the hippocampal mossy fiber synapse. Proceedings of the National Academy of Sciences of the United States of America, 105(33), 11998-2003. [PubMed:18687898] [PMC] [WorldCat] [DOI] - ↑
Kakizawa, S., Yamazawa, T., Chen, Y., Ito, A., Murayama, T., Oyamada, H., ..., & Iino, M. (2012).
Nitric oxide-induced calcium release via ryanodine receptors regulates neuronal function. The EMBO journal, 31(2), 417-28. [PubMed:22036948] [PMC] [WorldCat] [DOI] - ↑
Kakizawa, S., Moriguchi, S., Ikeda, A., Iino, M., & Takeshima, H. (2008).
Functional crosstalk between cell-surface and intracellular channels mediated by junctophilins essential for neuronal functions. Cerebellum (London, England), 7(3), 385-91. [PubMed:18607668] [WorldCat] [DOI]